

Dynamic limbic networks and social diversity in vertebrates: from neural context to neuromodulatory patterning
- PMID: 19520105
- PMCID: PMC2763925
- DOI: 10.1016/j.yfrne.2009.05.007
Dynamic limbic networks and social diversity in vertebrates: from neural context to neuromodulatory patterning
Abstract
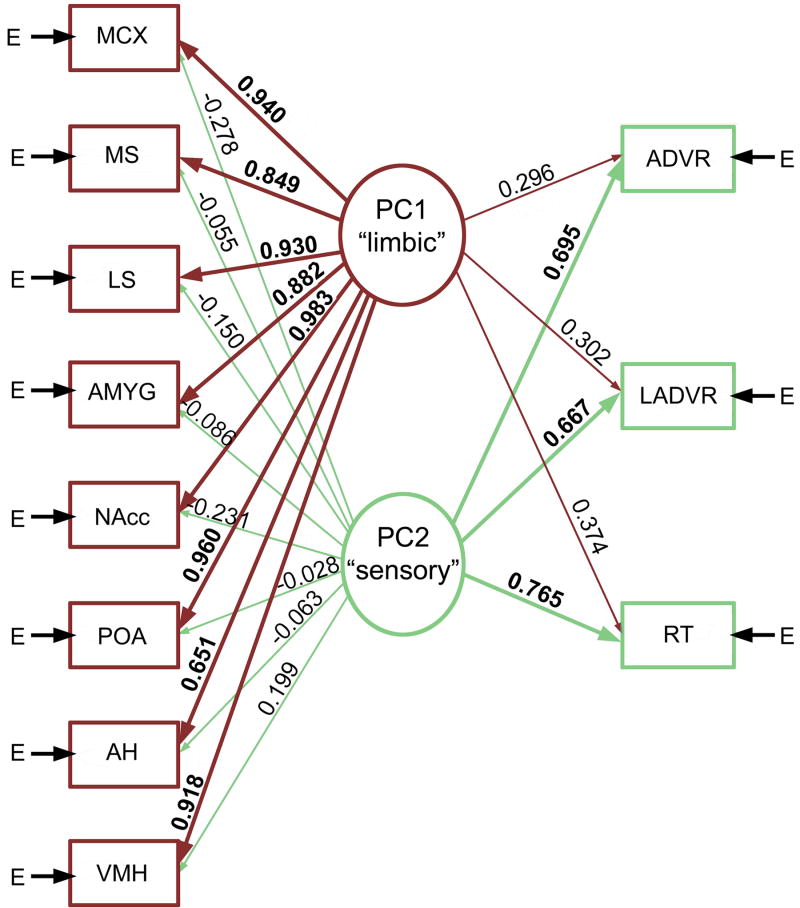
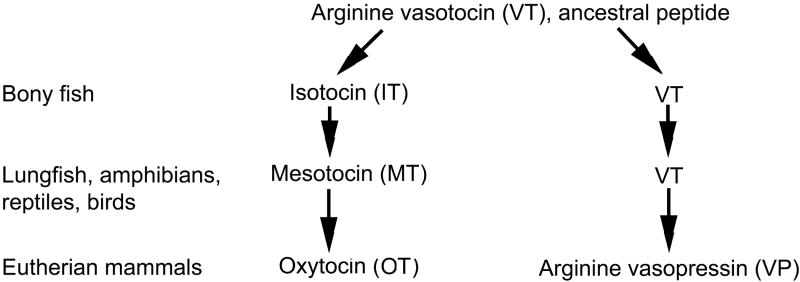
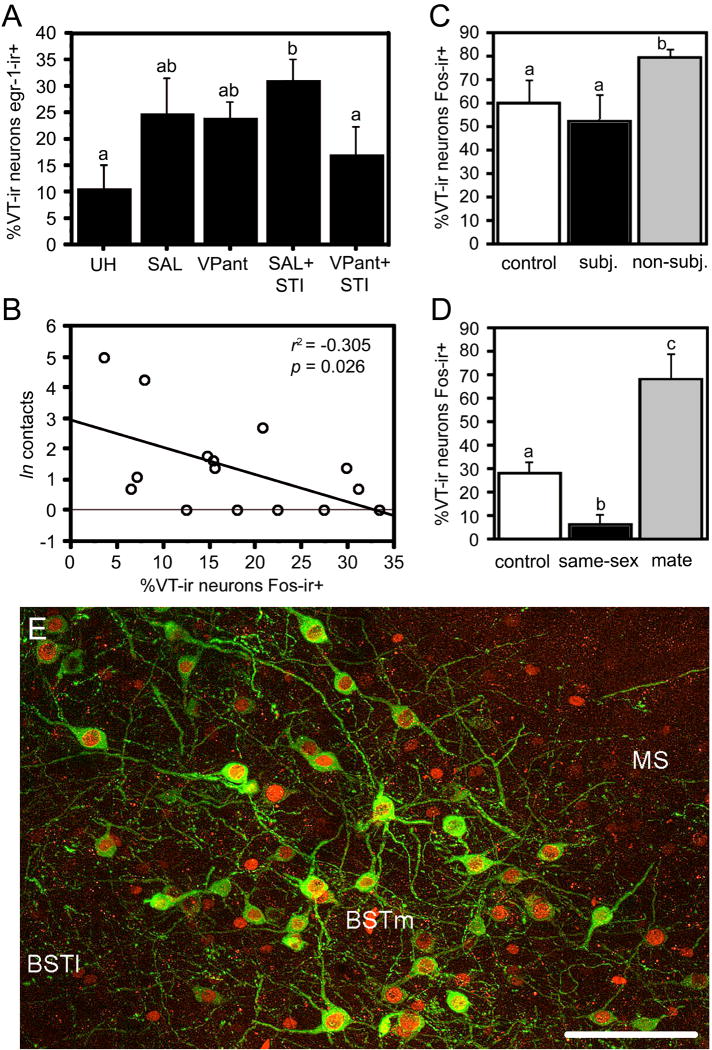
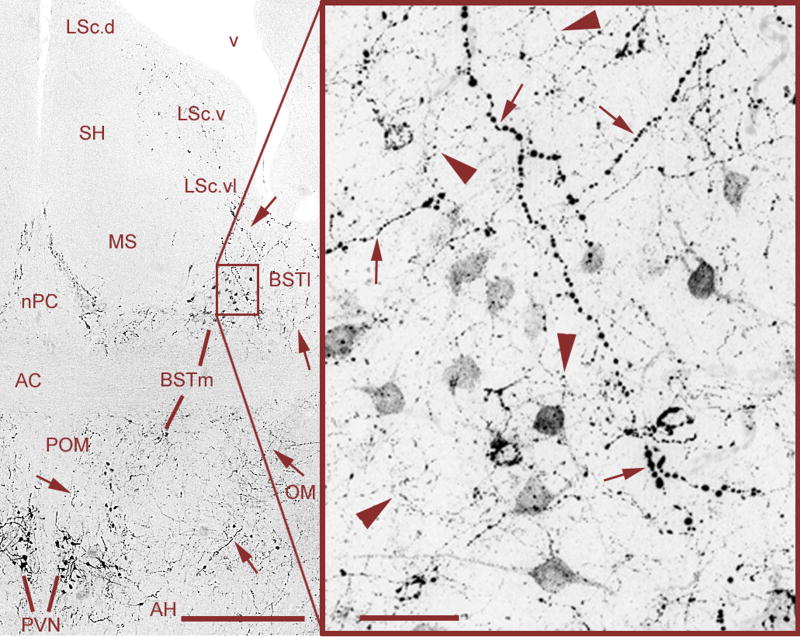
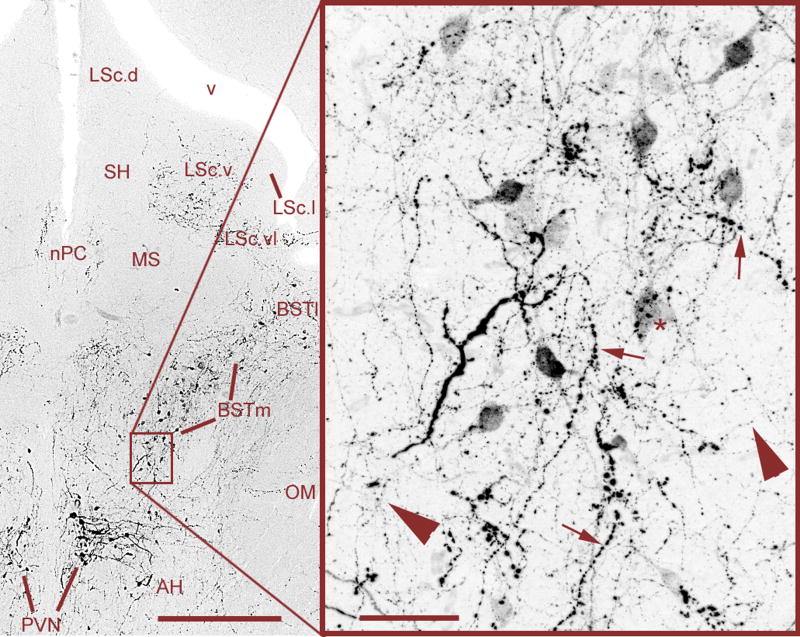
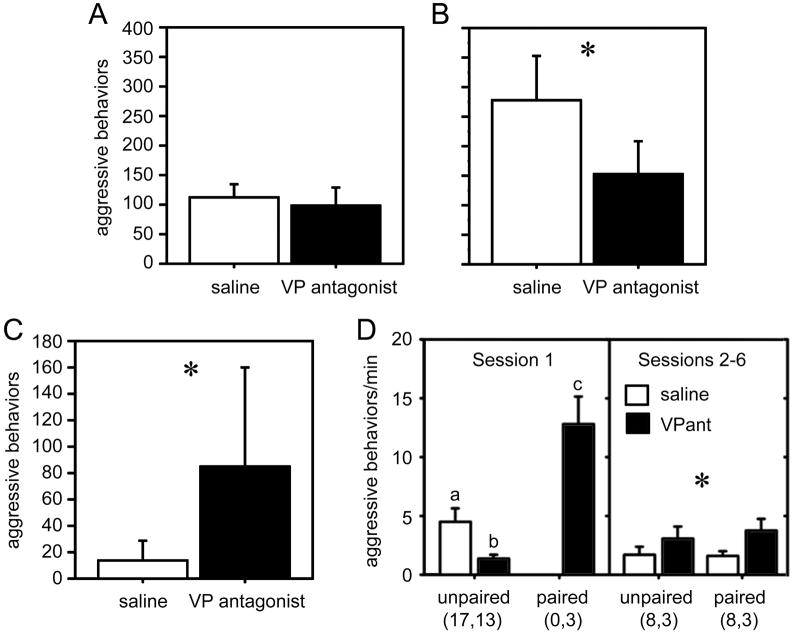
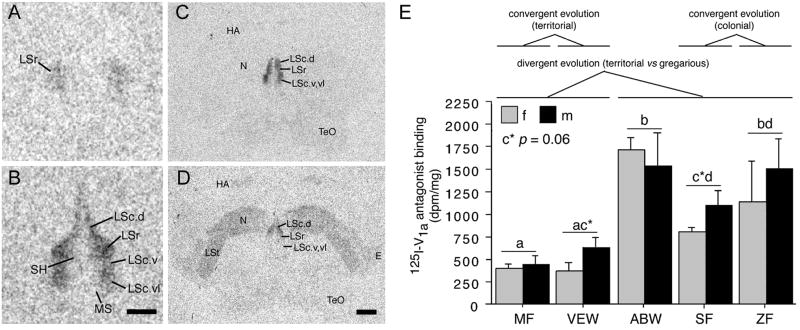
Vertebrate animals exhibit a spectacular diversity of social behaviors, yet a variety of basic social behavior processes are essential to all species. These include social signaling; discrimination of conspecifics and sexual partners; appetitive and consummatory sexual behaviors; aggression and dominance behaviors; and parental behaviors (the latter with rare exceptions). These behaviors are of fundamental importance and are regulated by an evolutionarily conserved, core social behavior network (SBN) of the limbic forebrain and midbrain. The SBN encodes social information in a highly dynamic, distributed manner, such that behavior is most strongly linked to the pattern of neural activity across the SBN, not the activity of single loci. Thus, shifts in the relative weighting of activity across SBN nodes can conceivably produce almost limitless variation in behavior, including diversity across species (as weighting is modified through evolution), across behavioral contexts (as weights change temporally) and across behavioral phenotypes (as weighting is specified through heritable and developmental processes). Individual neural loci may also express diverse relationships to behavior, depending upon temporal variations in their functional connectivity to other brain regions ("neural context"). We here review the basic properties of the SBN and show how behavioral variation relates to functional connectivity of the network, and discuss ways in which neuroendocrine factors adjust network activity to produce behavioral diversity. In addition to the actions of steroid hormones on SBN state, we examine the temporally plastic and evolutionarily labile properties of the nonapeptides (the vasopressin- and oxytocin-like neuropeptides), and show how variations in nonapeptide signaling within the SBN serve to promote behavioral diversity across social contexts, seasons, phenotypes and species. Although this diversity is daunting in its complexity, the search for common "organizing principles" has become increasingly fruitful. We focus on multiple aspects of behavior, including sexual behavior, aggression and affiliation, and in each of these areas, we show how broadly relevant insights have been obtained through the examination of behavioral diversity in a wide range of vertebrate taxa.
Figures
References
-
- Acher R. Chemistry of the neurohypophysial hormones: an example of molecular evolution. Endocrinology, American Physiological Society; Washington, D.C.: 1972. pp. 119–130.
-
- Adamantidis A, de Lecea L. Sleep and metabolism: shared circuits, new connections. Trends Endocrinol Metab. 2008;19:362–370. - PubMed
-
- Balfour ME, Brown JL, Yu L, Coolen LM. Potential contributions of efferents from medial prefrontal cortex to neural activation following sexual behavior in the male rat. Neuroscience. 2006;137:1259–1276. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
