

Fur controls iron homeostasis and oxidative stress defense in the oligotrophic alpha-proteobacterium Caulobacter crescentus
- PMID: 19520766
- PMCID: PMC2724300
- DOI: 10.1093/nar/gkp509
Fur controls iron homeostasis and oxidative stress defense in the oligotrophic alpha-proteobacterium Caulobacter crescentus
Abstract
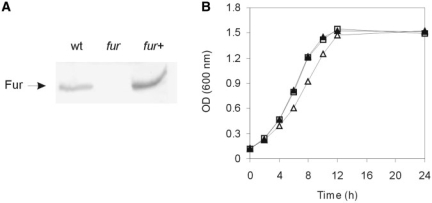
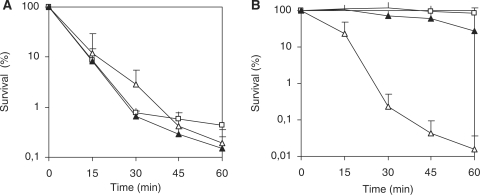
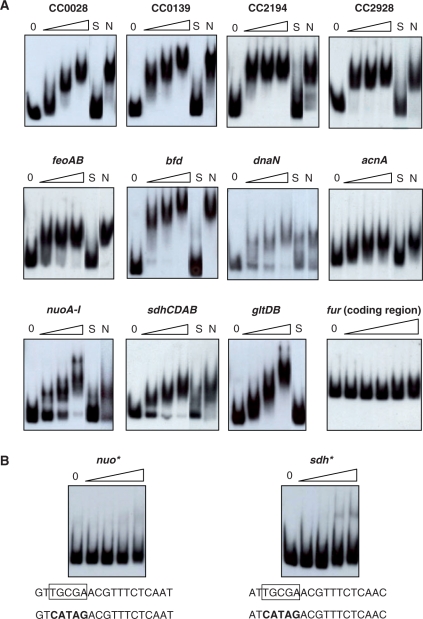
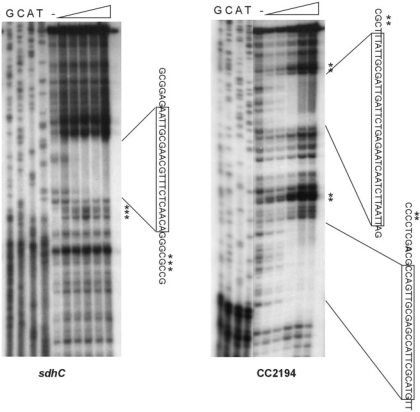
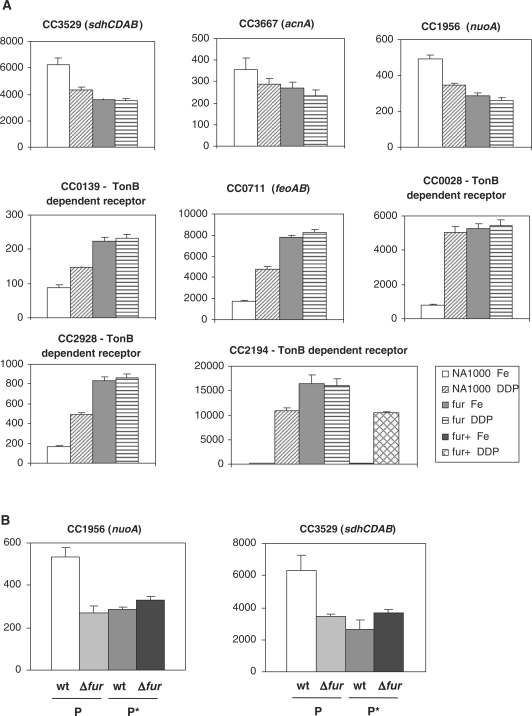
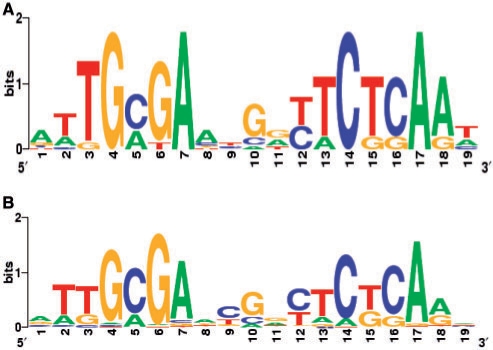
In most bacteria, the ferric uptake regulator (Fur) is a global regulator that controls iron homeostasis and other cellular processes, such as oxidative stress defense. In this work, we apply a combination of bioinformatics, in vitro and in vivo assays to identify the Caulobacter crescentus Fur regulon. A C. crescentus fur deletion mutant showed a slow growth phenotype, and was hypersensitive to H(2)O(2) and organic peroxide. Using a position weight matrix approach, several predicted Fur-binding sites were detected in the genome of C. crescentus, located in regulatory regions of genes not only involved in iron uptake and usage but also in other functions. Selected Fur-binding sites were validated using electrophoretic mobility shift assay and DNAse I footprinting analysis. Gene expression assays revealed that genes involved in iron uptake were repressed by iron-Fur and induced under conditions of iron limitation, whereas genes encoding iron-using proteins were activated by Fur under conditions of iron sufficiency. Furthermore, several genes that are regulated via small RNAs in other bacteria were found to be directly regulated by Fur in C. crescentus. In conclusion, Fur functions as an activator and as a repressor, integrating iron metabolism and oxidative stress response in C. crescentus.
Figures

Similar articles
-
Global transcriptional response of Caulobacter crescentus to iron availability.BMC Genomics. 2013 Aug 13;14:549. doi: 10.1186/1471-2164-14-549. BMC Genomics. 2013. PMID: 23941329 Free PMC article.
-
Cold Regulation of Genes Encoding Ion Transport Systems in the Oligotrophic Bacterium Caulobacter crescentus.Microbiol Spectr. 2021 Sep 3;9(1):e0071021. doi: 10.1128/Spectrum.00710-21. Epub 2021 Aug 25. Microbiol Spectr. 2021. PMID: 34479415 Free PMC article.
-
Role and regulation of ferritin-like proteins in iron homeostasis and oxidative stress survival of Caulobacter crescentus.Biometals. 2016 Oct;29(5):851-62. doi: 10.1007/s10534-016-9956-y. Epub 2016 Aug 2. Biometals. 2016. PMID: 27484774
-
Transcriptional regulation by Ferric Uptake Regulator (Fur) in pathogenic bacteria.Front Cell Infect Microbiol. 2013 Oct 2;3:59. doi: 10.3389/fcimb.2013.00059. eCollection 2013. Front Cell Infect Microbiol. 2013. PMID: 24106689 Free PMC article. Review.
-
How we learnt about iron acquisition in Pseudomonas aeruginosa: a series of very fortunate events.Biometals. 2007 Jun;20(3-4):587-601. doi: 10.1007/s10534-006-9067-2. Epub 2006 Dec 22. Biometals. 2007. PMID: 17186376 Review.
Cited by
-
The Regulatory Protein ChuP Connects Heme and Siderophore-Mediated Iron Acquisition Systems Required for Chromobacterium violaceum Virulence.Front Cell Infect Microbiol. 2022 May 11;12:873536. doi: 10.3389/fcimb.2022.873536. eCollection 2022. Front Cell Infect Microbiol. 2022. PMID: 35646721 Free PMC article.
-
The Caulobacter NtrB-NtrC two-component system bridges nitrogen assimilation and cell development.bioRxiv [Preprint]. 2023 Aug 31:2023.06.06.543975. doi: 10.1101/2023.06.06.543975. bioRxiv. 2023. Update in: J Bacteriol. 2023 Oct 26;205(10):e0018123. doi: 10.1128/jb.00181-23. PMID: 37333394 Free PMC article. Updated. Preprint.
-
New evolving strategies revealed by transcriptomic analysis of a fur- mutant of the cyanotrophic bacterium Pseudomonas pseudoalcaligenes CECT 5344.Microb Biotechnol. 2020 Jan;13(1):148-161. doi: 10.1111/1751-7915.13408. Epub 2019 Apr 21. Microb Biotechnol. 2020. PMID: 31006999 Free PMC article.
-
Analysis of the Caulobacter crescentus Zur regulon reveals novel insights in zinc acquisition by TonB-dependent outer membrane proteins.BMC Genomics. 2014 Aug 28;15(1):734. doi: 10.1186/1471-2164-15-734. BMC Genomics. 2014. PMID: 25168179 Free PMC article.
-
Functional characterization of RhuB as a second TonB2-dependent hemin receptor in Riemerella anatipestifer CH-1.Microbiol Spectr. 2024 Apr 2;12(4):e0313323. doi: 10.1128/spectrum.03133-23. Epub 2024 Feb 20. Microbiol Spectr. 2024. PMID: 38376226 Free PMC article.
References
-
- Andrews SC, Robinson AK, Rodriguez-Quinones F. Bacterial iron homeostasis. FEMS Microbiol. Rev. 2003;27:215–237. - PubMed
-
- Schaible UE, Kaufmann SHE. Iron and microbial infection. Nat. Rev. Microbiol. 2004;2:946–953. - PubMed
-
- Wandersman C, Delepelaire P. Bacterial iron sources: from siderophores to hemophores. Annu. Rev. Microbiol. 2004;58:611–647. - PubMed
-
- Braun V, Braun M. Active transport of iron and siderophore antibiotics. Curr. Opin. Microbiol. 2002;5:194–201. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
