

Zinc coordination is required for and regulates transcription activation by Epstein-Barr nuclear antigen 1
- PMID: 19521517
- PMCID: PMC2690687
- DOI: 10.1371/journal.ppat.1000469
Zinc coordination is required for and regulates transcription activation by Epstein-Barr nuclear antigen 1
Abstract
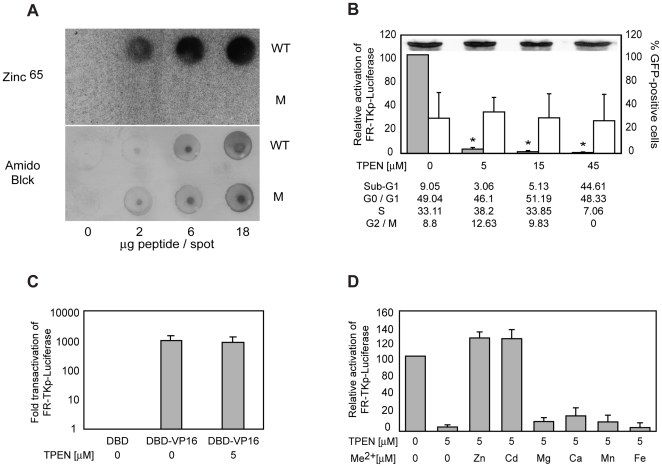
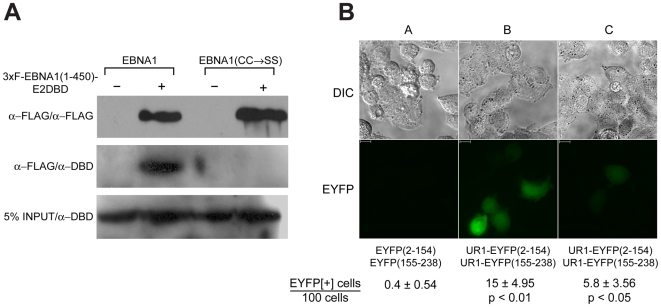
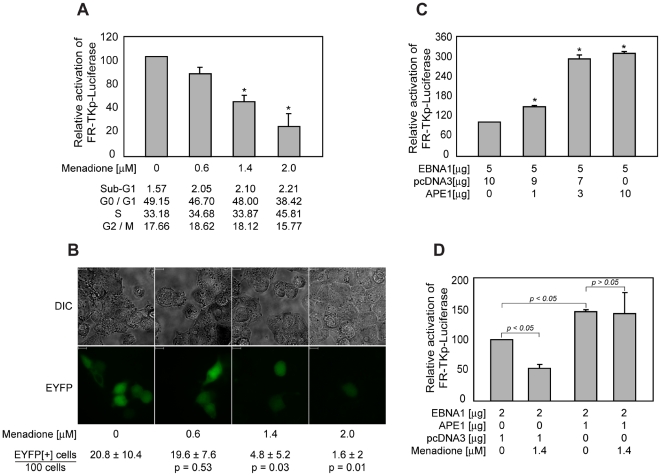
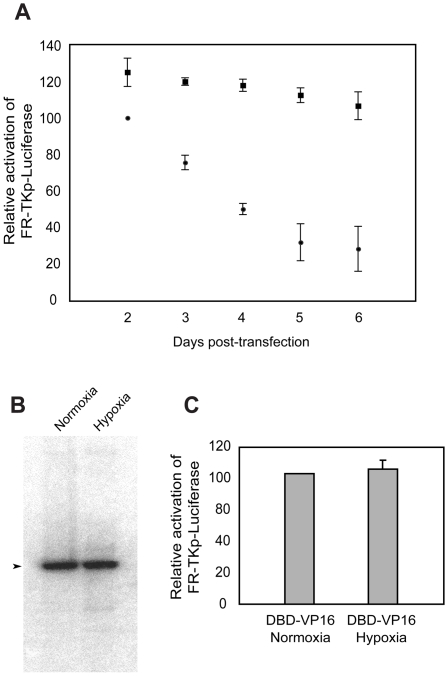
Epstein-Barr Nuclear Antigen 1 (EBNA1) is essential for Epstein-Barr virus to immortalize naïve B-cells. Upon binding a cluster of 20 cognate binding-sites termed the family of repeats, EBNA1 transactivates promoters for EBV genes that are required for immortalization. A small domain, termed UR1, that is 25 amino-acids in length, has been identified previously as essential for EBNA1 to activate transcription. In this study, we have elucidated how UR1 contributes to EBNA1's ability to transactivate. We show that zinc is necessary for EBNA1 to activate transcription, and that UR1 coordinates zinc through a pair of essential cysteines contained within it. UR1 dimerizes upon coordinating zinc, indicating that EBNA1 contains a second dimerization interface in its amino-terminus. There is a strong correlation between UR1-mediated dimerization and EBNA1's ability to transactivate cooperatively. Point mutants of EBNA1 that disrupt zinc coordination also prevent self-association, and do not activate transcription cooperatively. Further, we demonstrate that UR1 acts as a molecular sensor that regulates the ability of EBNA1 to activate transcription in response to changes in redox and oxygen partial pressure (pO(2)). Mild oxidative stress mimicking such environmental changes decreases EBNA1-dependent transcription in a lymphoblastoid cell-line. Coincident with a reduction in EBNA1-dependent transcription, reductions are observed in EBNA2 and LMP1 protein levels. Although these changes do not affect LCL survival, treated cells accumulate in G0/G1. These findings are discussed in the context of EBV latency in body compartments that differ strikingly in their pO(2) and redox potential.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures




References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
