

Coordinated surface activities in Variovorax paradoxus EPS
- PMID: 19523213
- PMCID: PMC2704215
- DOI: 10.1186/1471-2180-9-124
Coordinated surface activities in Variovorax paradoxus EPS
Abstract
Background: Variovorax paradoxus is an aerobic soil bacterium frequently associated with important biodegradative processes in nature. Our group has cultivated a mucoid strain of Variovorax paradoxus for study as a model of bacterial development and response to environmental conditions. Colonies of this organism vary widely in appearance depending on agar plate type.
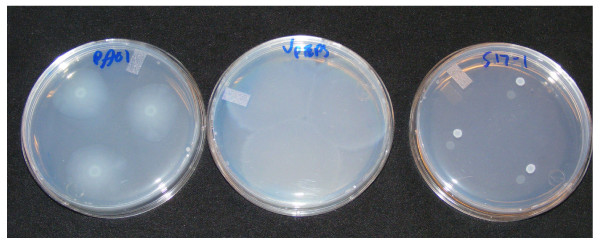
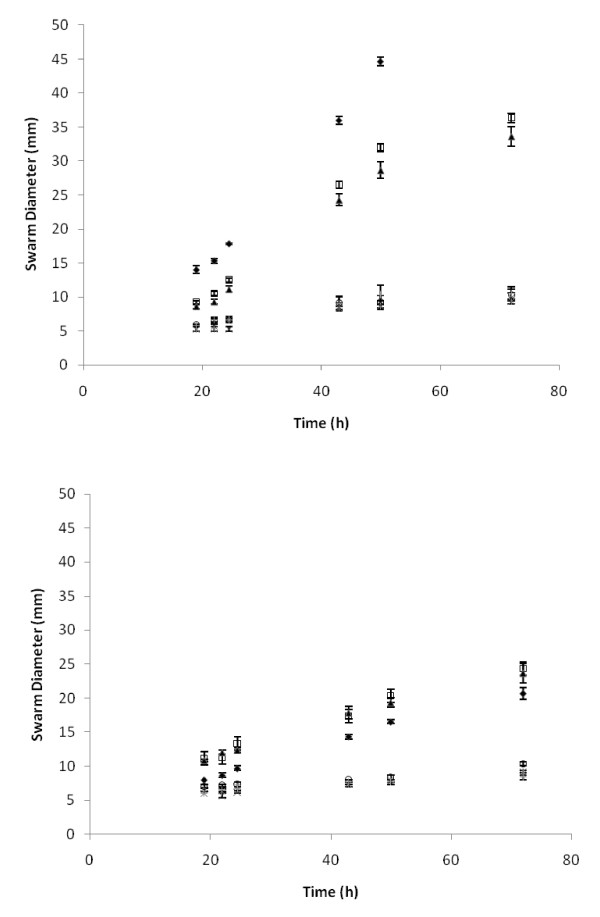
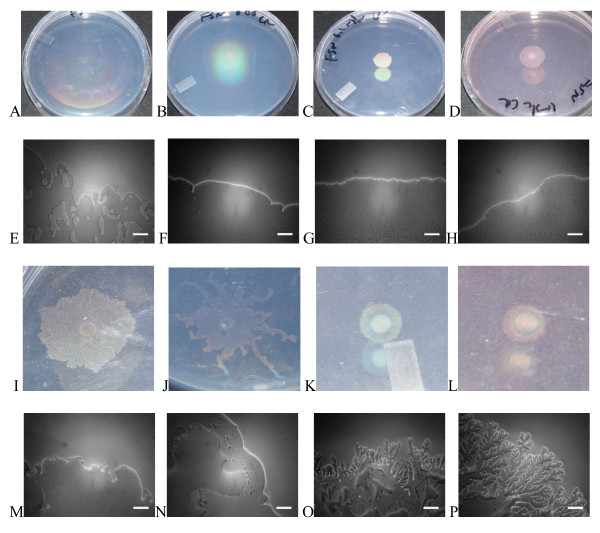
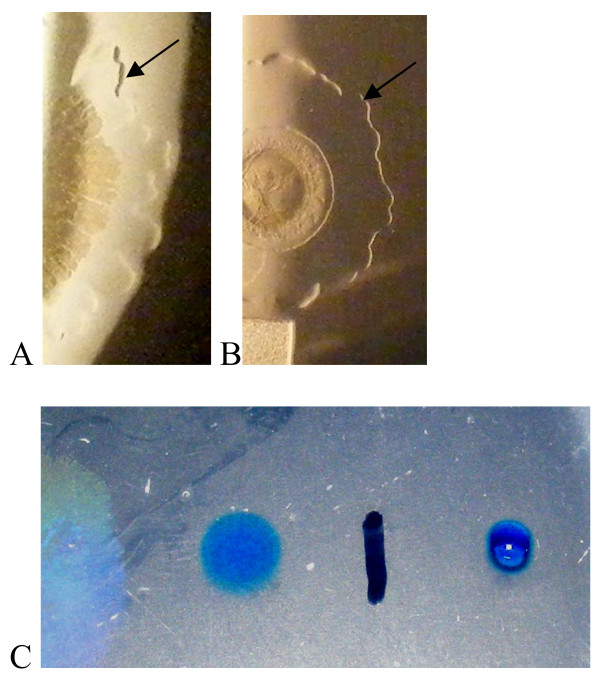
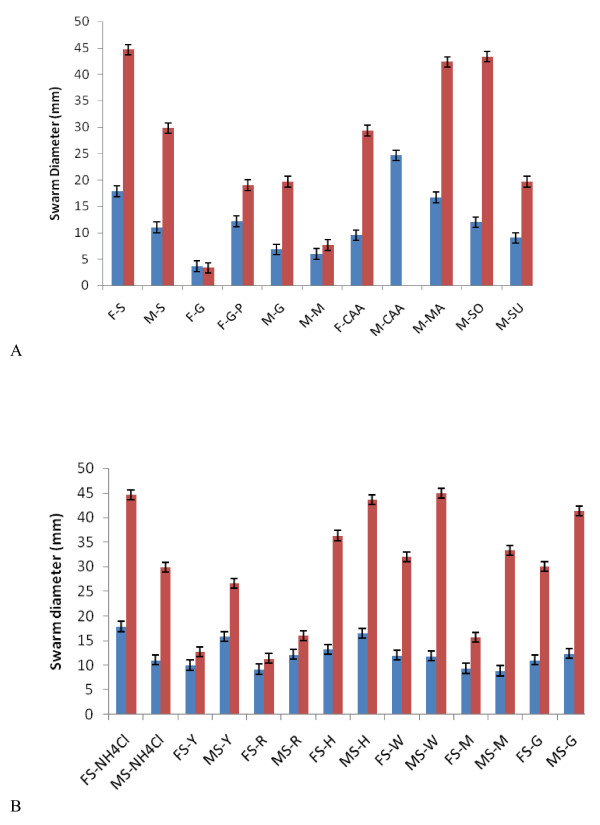
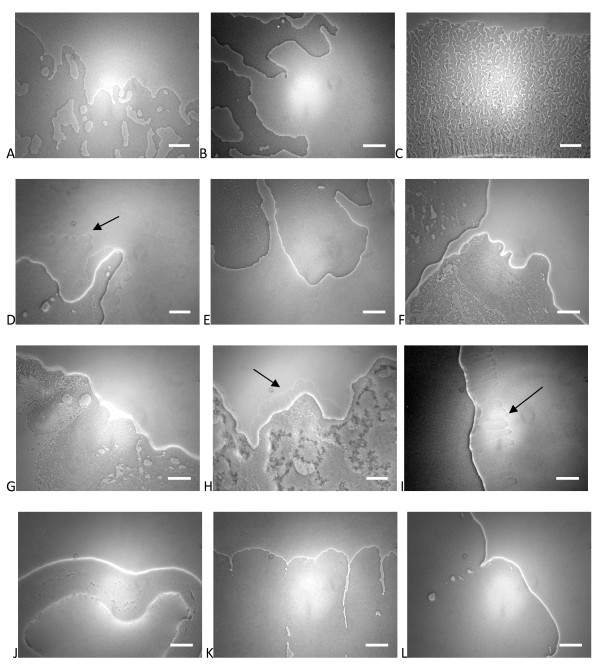
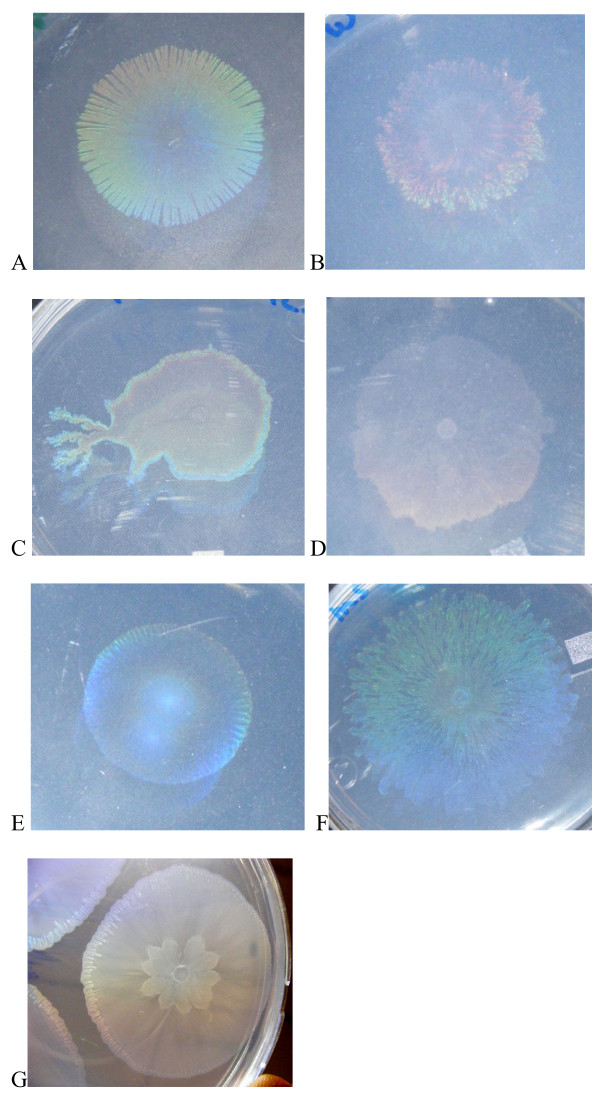
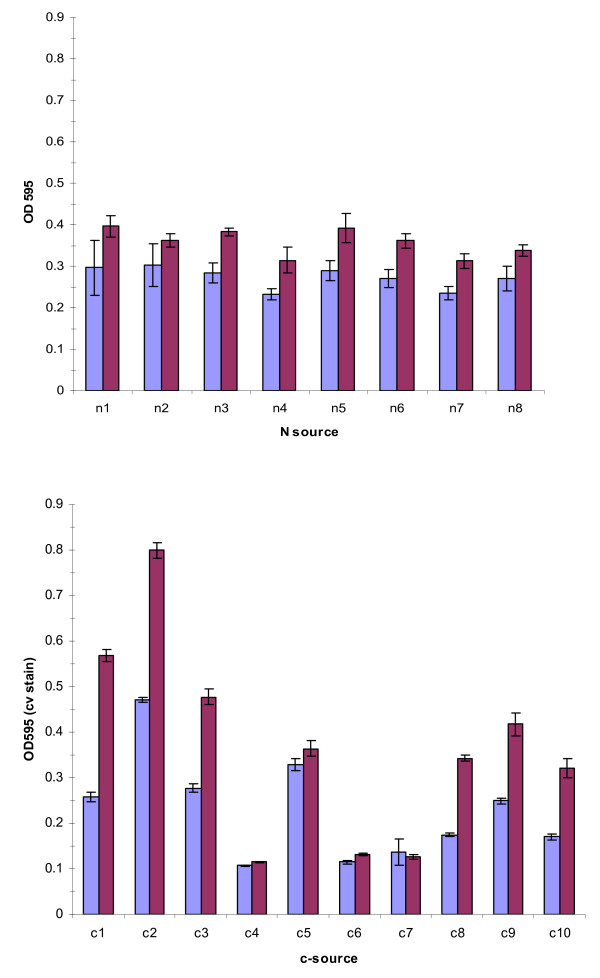
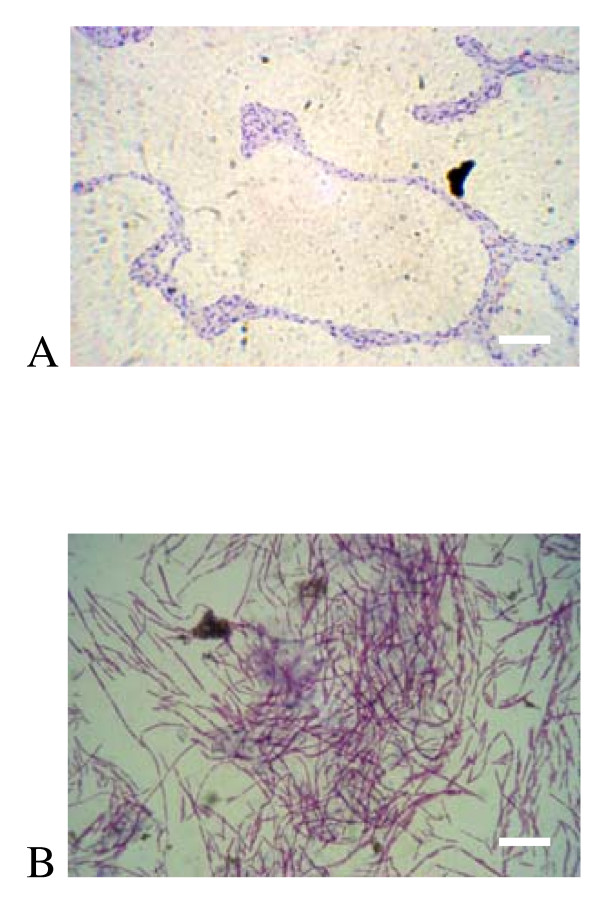
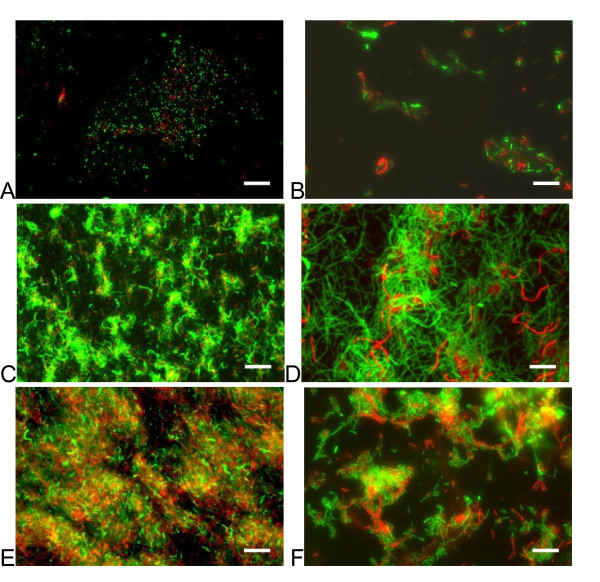
Results: Surface motility was observed on minimal defined agar plates with 0.5% agarose, similar in nature to swarming motility identified in Pseudomonas aeruginosa PAO1. We examined this motility under several culture conditions, including inhibition of flagellar motility using Congo Red. We demonstrated that the presence of a wetting agent, mineral, and nutrient content of the media altered the swarming phenotype. We also demonstrated that the wetting agent reduces the surface tension of the agar. We were able to directly observe the presence of the wetting agent in the presence and absence of Congo Red, and found that incubation in a humidified chamber inhibited the production of wetting agent, and also slowed the progression of the swarming colony. We observed that swarming was related to both carbon and nitrogen sources, as well as mineral salts base. The phosphate concentration of the mineral base was critical for growth and swarming on glucose, but not succinate. Swarming on other carbon sources was generally only observed using M9 salts mineral base. Rapid swarming was observed on malic acid, d-sorbitol, casamino acids, and succinate. Swarming at a lower but still detectable rate was observed on glucose and sucrose, with weak swarming on maltose. Nitrogen source tests using succinate as carbon source demonstrated two distinct forms of swarming, with very different macroscopic swarm characteristics. Rapid swarming was observed when ammonium ion was provided as nitrogen source, as well as when histidine, tryptophan, or glycine was provided. Slower swarming was observed with methionine, arginine, or tyrosine. Large effects of mineral content on swarming were seen with tyrosine and methionine as nitrogen sources. Biofilms form readily under various culture circumstances, and show wide variance in structure under different conditions. The amount of biofilm as measured by crystal violet retention was dependent on carbon source, but not nitrogen source. Filamentous growth in the biofilm depends on shear stress, and is enhanced by continuous input of nutrients in chemostat culture.
Conclusion: Our studies have established that the beta-proteobacterium Variovorax paradoxus displays a number of distinct physiologies when grown on surfaces, indicative of a complex response to several growth parameters. We have identified a number of factors that drive sessile and motile surface phenotypes. This work forms a basis for future studies using this genetically tractable soil bacterium to study the regulation of microbial development on surfaces.
Figures
Similar articles
-
Genes that influence swarming motility and biofilm formation in Variovorax paradoxus EPS.PLoS One. 2012;7(2):e31832. doi: 10.1371/journal.pone.0031832. Epub 2012 Feb 21. PLoS One. 2012. PMID: 22363744 Free PMC article.
-
A novel semi-selective medium for Pseudomonas protegens isolation from soil samples.J Microbiol Methods. 2020 May;172:105911. doi: 10.1016/j.mimet.2020.105911. Epub 2020 Mar 30. J Microbiol Methods. 2020. PMID: 32240707
-
Swarming and complex pattern formation in Paenibacillus vortex studied by imaging and tracking cells.BMC Microbiol. 2008 Feb 25;8:36. doi: 10.1186/1471-2180-8-36. BMC Microbiol. 2008. PMID: 18298829 Free PMC article.
-
Metabolic characteristics of the species Variovorax paradoxus.Appl Microbiol Biotechnol. 2013 Jan;97(2):541-60. doi: 10.1007/s00253-012-4585-z. Epub 2012 Nov 29. Appl Microbiol Biotechnol. 2013. PMID: 23192768 Review.
-
[Characterization of bacterial swarming motility: a review].Sheng Wu Gong Cheng Xue Bao. 2023 Aug 25;39(8):3188-3203. doi: 10.13345/j.cjb.220892. Sheng Wu Gong Cheng Xue Bao. 2023. PMID: 37622355 Review. Chinese.
Cited by
-
Genes that influence swarming motility and biofilm formation in Variovorax paradoxus EPS.PLoS One. 2012;7(2):e31832. doi: 10.1371/journal.pone.0031832. Epub 2012 Feb 21. PLoS One. 2012. PMID: 22363744 Free PMC article.
-
Phenanthrene Degradation by Photosynthetic Bacterial Consortium Dominated by Fischerella sp.Life (Basel). 2023 Apr 28;13(5):1108. doi: 10.3390/life13051108. Life (Basel). 2023. PMID: 37240753 Free PMC article.
-
Succinyl-CoA:3-sulfinopropionate CoA-transferase from Variovorax paradoxus strain TBEA6, a novel member of the class III coenzyme A (CoA)-transferase family.J Bacteriol. 2013 Aug;195(16):3761-73. doi: 10.1128/JB.00456-13. Epub 2013 Jun 14. J Bacteriol. 2013. PMID: 23772073 Free PMC article.
-
Biofilm formation by Psychrobacter arcticus and the role of a large adhesin in attachment to surfaces.Appl Environ Microbiol. 2013 Jul;79(13):3967-73. doi: 10.1128/AEM.00867-13. Epub 2013 Apr 19. Appl Environ Microbiol. 2013. PMID: 23603675 Free PMC article.
-
Surveying a Swarm: Experimental Techniques To Establish and Examine Bacterial Collective Motion.Appl Environ Microbiol. 2022 Feb 8;88(3):e0185321. doi: 10.1128/AEM.01853-21. Epub 2021 Dec 8. Appl Environ Microbiol. 2022. PMID: 34878816 Free PMC article.
References
-
- Willems A, Ley JD, Gillis M, Kersters K. Comamonadaceae, a new family encompassing the Acidovorans rRNA complex, including Variovorax paradoxus gen. nov., comb. nov., for Alcaligenes paradoxus (Davis 1969) Int J Syst Bacteriol. 1991;41(445–450)
-
- Trusova MY, Gladyshev MI. Phylogenetic diversity of winter bacterioplankton of eutrophic siberian reservoirs as revealed by 16S rRNA gene sequence. Microb Ecol. 2002;44(3):252–259. - PubMed
-
- Smith D, Alvey S, Crowley DE. Cooperative catabolic pathways within an atrazine-degrading enrichment culture isolated from soil. FEMS Microbiol Ecol. 2005;53(2):265–273. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
