

An unfolded CH1 domain controls the assembly and secretion of IgG antibodies
- PMID: 19524537
- PMCID: PMC2908990
- DOI: 10.1016/j.molcel.2009.04.028
An unfolded CH1 domain controls the assembly and secretion of IgG antibodies
Abstract
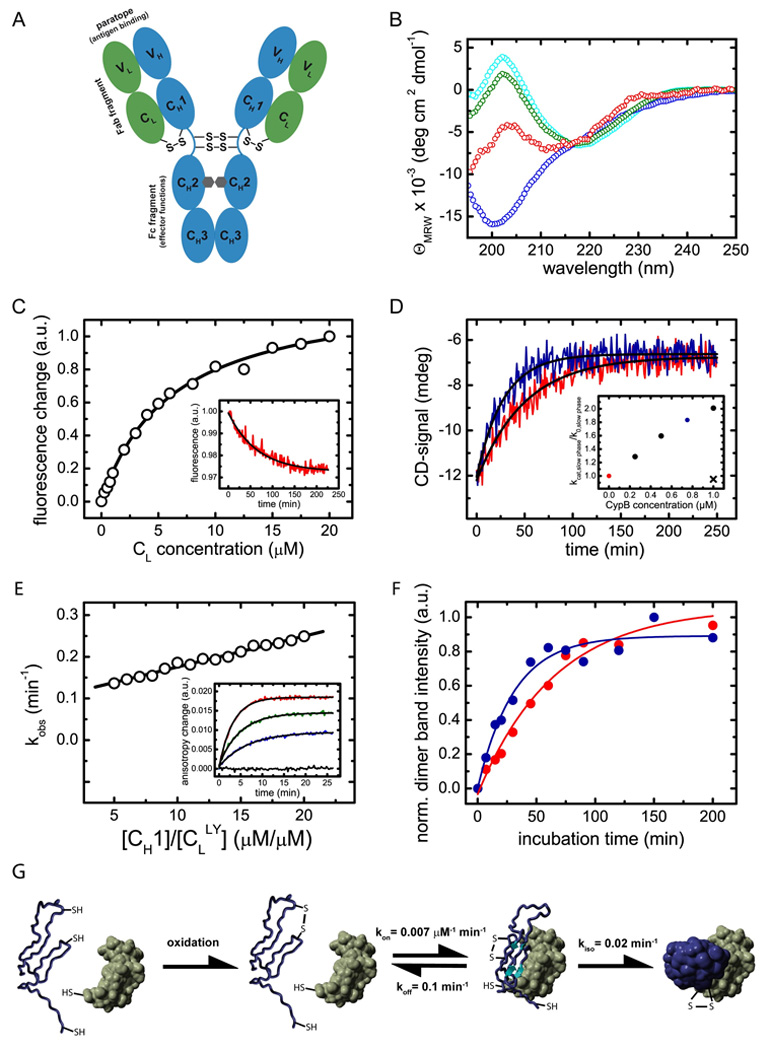
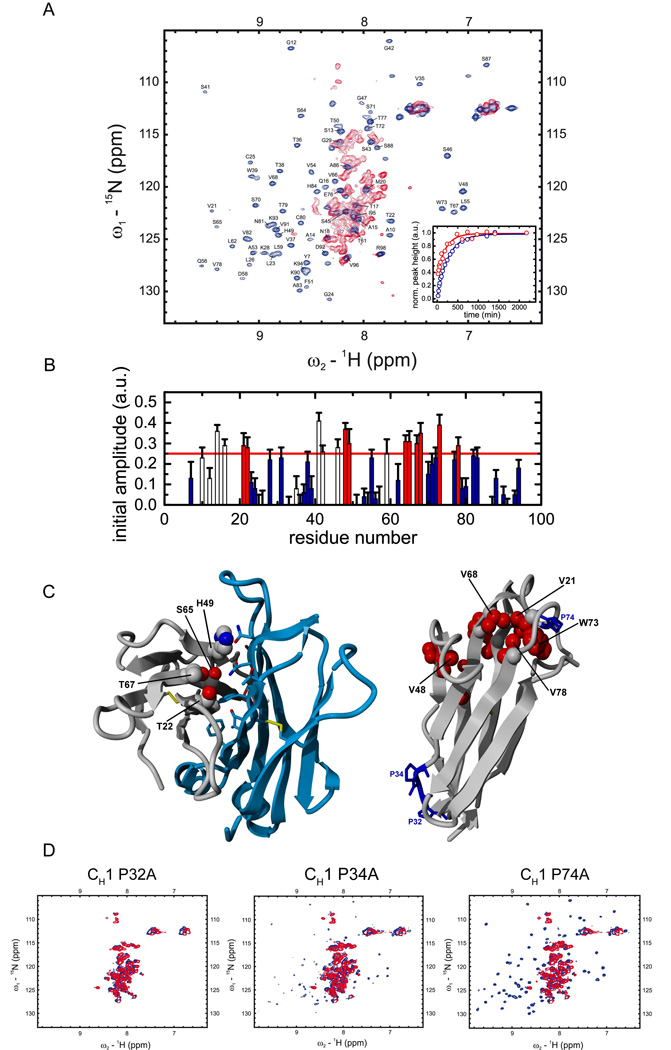
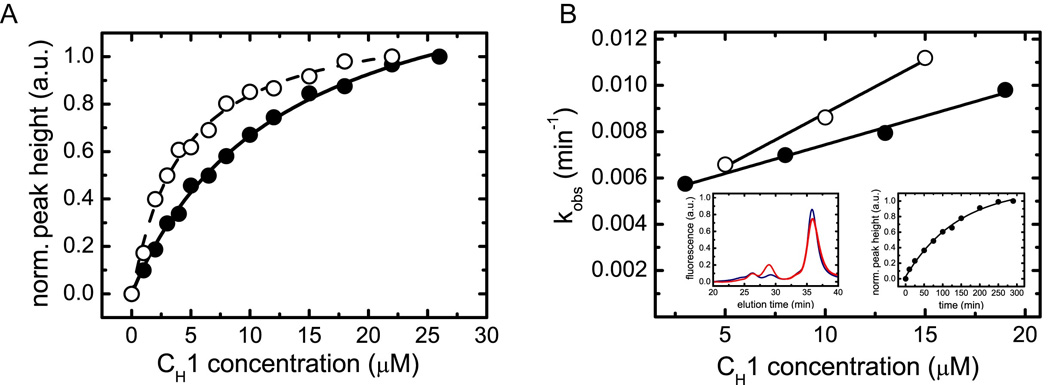
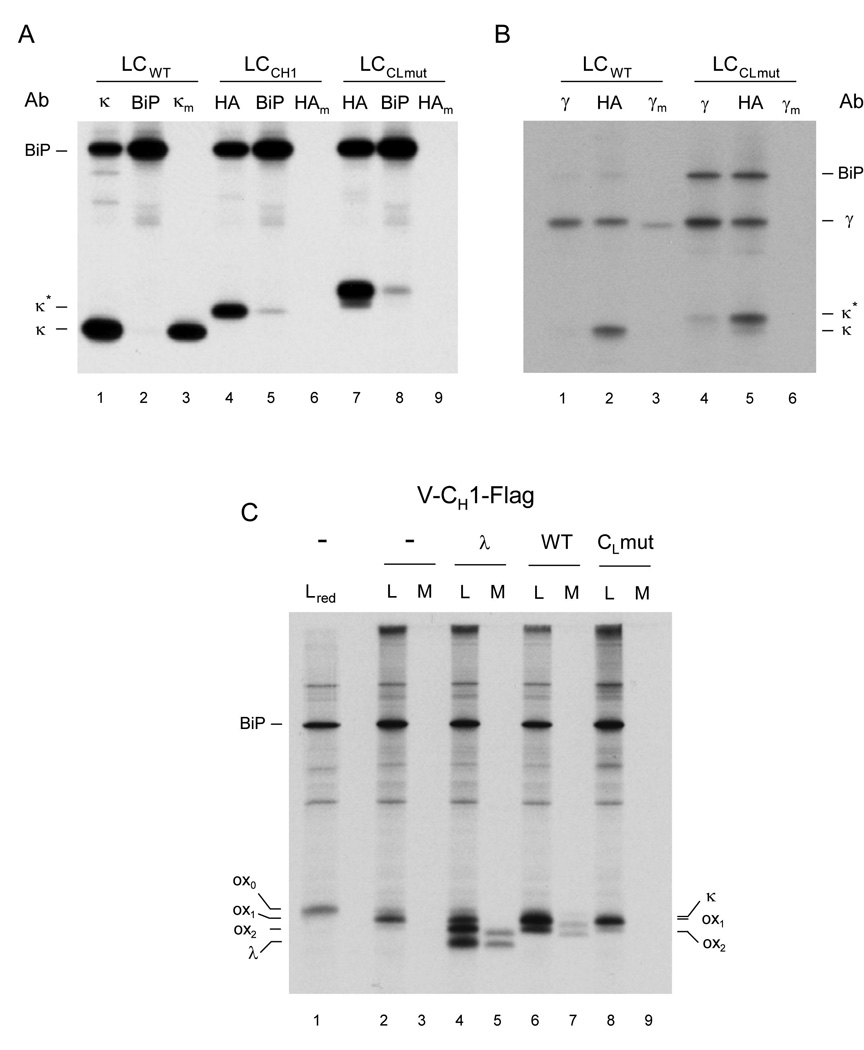
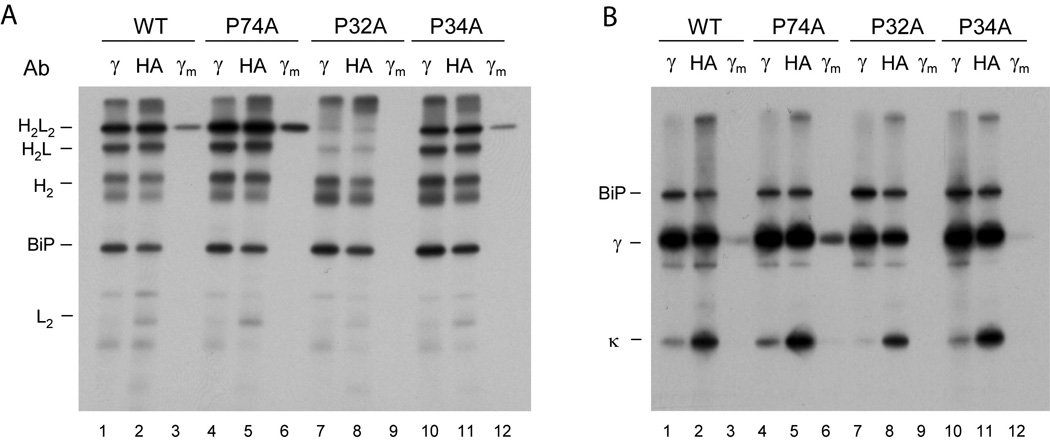
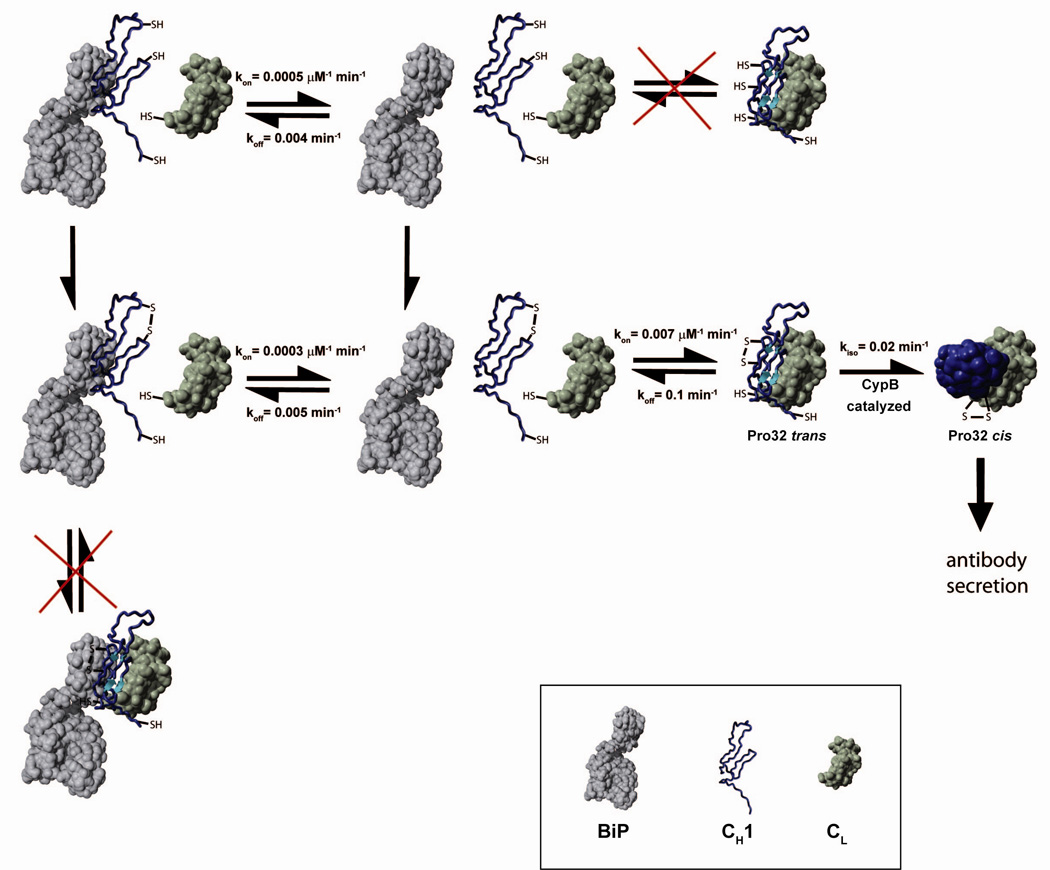
A prerequisite for antibody secretion and function is their assembly into a defined quaternary structure, composed of two heavy and two light chains for IgG. Unassembled heavy chains are actively retained in the endoplasmic reticulum (ER). Here, we show that the C(H)1 domain of the heavy chain is intrinsically disordered in vitro, which sets it apart from other antibody domains. It folds only upon interaction with the light-chain C(L) domain. Structure formation proceeds via a trapped intermediate and can be accelerated by the ER-specific peptidyl-prolyl isomerase cyclophilin B. The molecular chaperone BiP recognizes incompletely folded states of the C(H)1 domain and competes for binding to the C(L) domain. In vivo experiments demonstrate that requirements identified for folding the C(H)1 domain in vitro, including association with a folded C(L) domain and isomerization of a conserved proline residue, are essential for antibody assembly and secretion in the cell.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
Comment in
-
The molecular dating game: an antibody heavy chain hangs loose with a chaperone while waiting for its life partner.Mol Cell. 2009 Jun 26;34(6):635-6. doi: 10.1016/j.molcel.2009.06.006. Mol Cell. 2009. PMID: 19560414 Free PMC article.
References
-
- Adetugbo K. Spontaneous somatic mutations. Structural studies on mutant immunoglobulins. J. Biol. Chem. 1978;253:6076–6080. - PubMed
-
- Amzel LM, Poljak RJ. 3-Dimensional Structure of Immunoglobulins. Annual Review of Biochemistry. 1979;48:961–997. - PubMed
-
- Aricescu AR, Jones EY. Immunoglobulin superfamily cell adhesion molecules: zippers and signals. Curr. Opin. Cell Biol. 2007;19:543–550. - PubMed
-
- Augustine JG, de la Calle A, Knarr G, Buchner J, Frederick CA. The crystal structure of the Fab fragment of the monoclonal antibody MAK33 -Implications for folding and interaction with the chaperone BiP. Journal of Biological Chemistry. 2001;276:3287–3294. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
