

Adult birdsong is actively maintained by error correction
- PMID: 19525945
- PMCID: PMC2701972
- DOI: 10.1038/nn.2336
Adult birdsong is actively maintained by error correction
Abstract
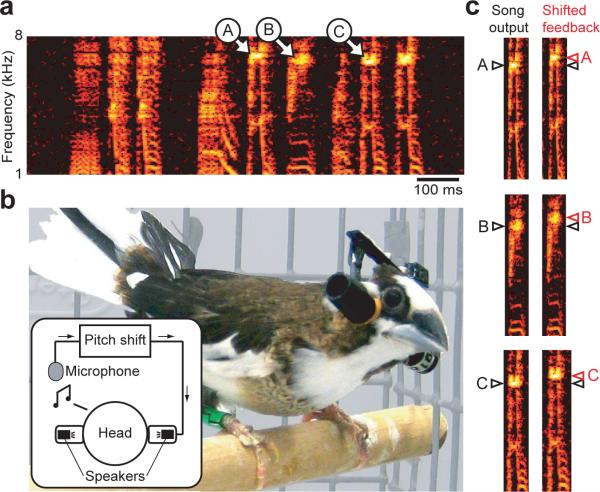
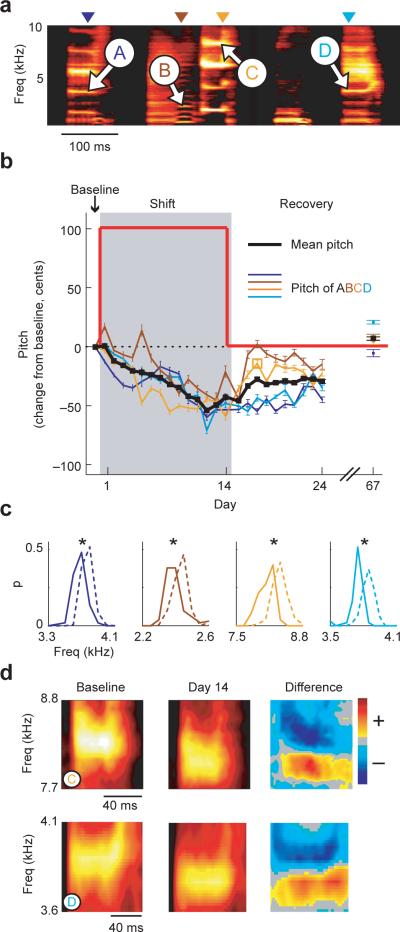
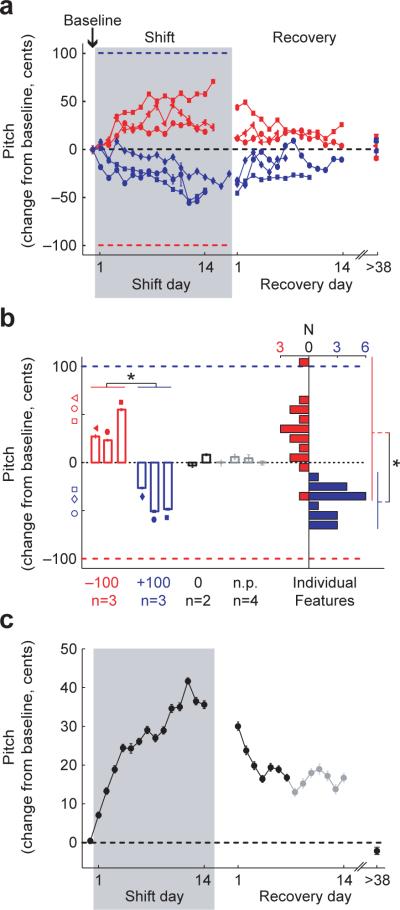
Humans learn to speak by a process of vocal imitation that requires the availability of auditory feedback. Similarly, young birds rely on auditory feedback when learning to imitate the songs of adult birds, providing one of the few examples of nonhuman vocal learning. However, although humans continue to use auditory feedback to correct vocal errors in adulthood, the mechanisms underlying the stability of adult birdsong are unknown. We found that, similar to human speech, adult birdsong is maintained by error correction. We perturbed the pitch (fundamental frequency) of auditory feedback in adult Bengalese finches using custom-designed headphones. Birds compensated for the imposed auditory error by adjusting the pitch of song. When the perturbation was removed, pitch returned to baseline. Our results indicate that adult birds correct vocal errors by comparing auditory feedback to a sensory target and suggest that lifelong error correction is a general principle of learned vocal behavior.
Figures
References
-
- Konishi M. The role of auditory feedback in the control of vocalization in the white-crowned sparrow. Zeitschrift fur Tierpsychologie. 1965;22:770–783. - PubMed
-
- Kuhl PK. Learning and representation in speech and language. Curr. Opin. in Neurobiol. 1995;4:812–822. - PubMed
-
- Arnold AP. The effects of castration on song development in zebra finches (Poephila guttata). J. Exp. Zool. 1975;191:261–278. - PubMed
-
- Kao MH, Doupe AJ, Brainard MS. Contributions of an avian basal gangliaforebrain circuit to real-time modulation of song. Nature. 2005;433:638–43. - PubMed
