

Attraction and activation of dendritic cells at the site of tumor elicits potent antitumor immunity
- PMID: 19532135
- PMCID: PMC2835257
- DOI: 10.1038/mt.2009.111
Attraction and activation of dendritic cells at the site of tumor elicits potent antitumor immunity
Abstract
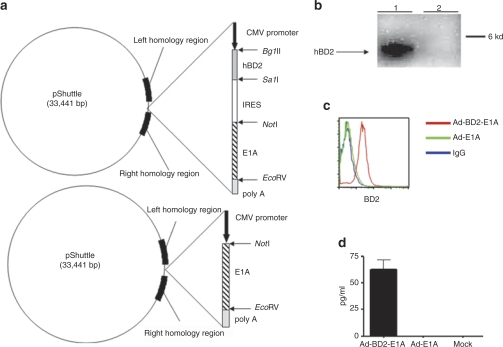
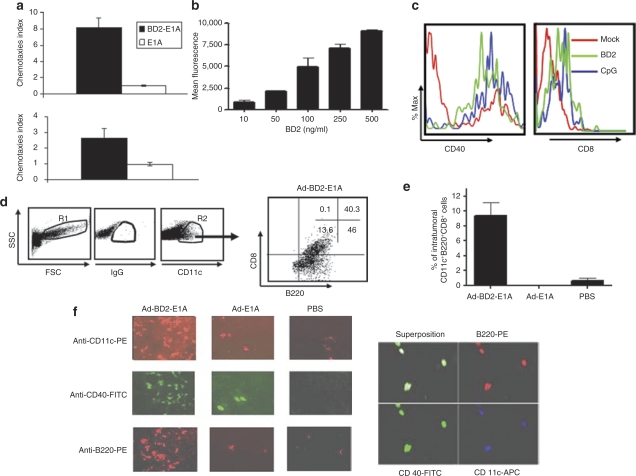
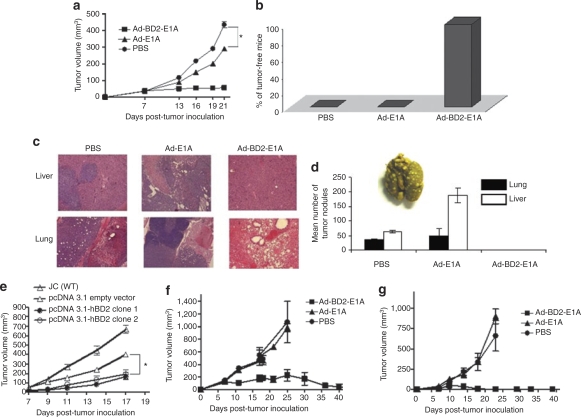
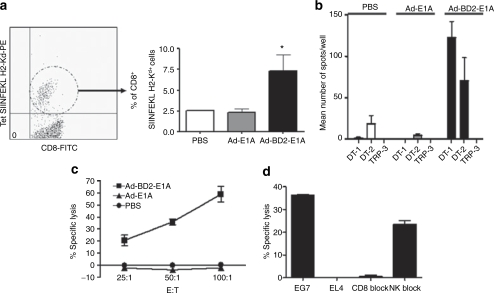
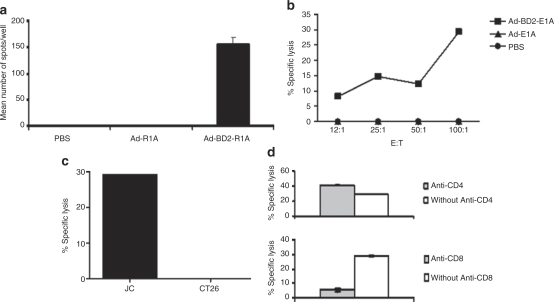
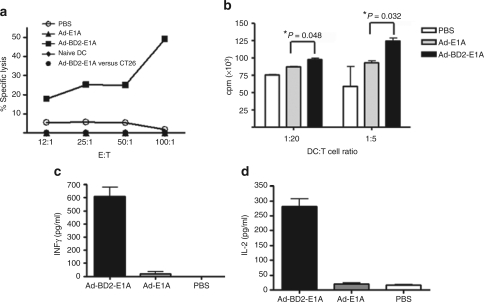
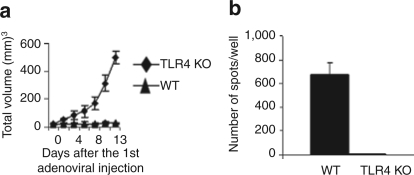
Tumor cells harbor unique genetic mutations, which lead to the generation of immunologically foreign antigenic peptide repertoire with the potential to induce individual tumor-specific immune responses. Here, we developed an in situ tumor vaccine with the ability to elicit antitumor immunity. This vaccine comprised an E1B-deleted oncolytic adenovirus expressing beta-defensin-2 (Ad-BD2-E1A) for releasing tumor antigens, recruiting and activating plasmacytoid dendritic cells (pDCs). Intratumoral injections of Ad-BD2-E1A vaccine inhibited primary breast tumor growth and blocked naturally occurring metastasis in mice. Ad-BD2-E1A vaccination induced potent tumor-specific T-cell responses. Splenic and intratumoral DCs isolated from Ad-BD2-E1A-immunized mice were able to stimulate or promote the differentiation of naive T cells into tumor-specific cytotoxic T cells. We further found that the increased numbers of mature CD45RA(+)CD8alpha(+)CD40(+) pDCs infiltrated into Ad-BD2-E1A-treated tumors. The antitumor effect of Ad-BD2-E1A vaccination was abrogated in toll-like receptor 4 (TLR4) deficient mice, suggesting the critical role of TLR4 in the induction of antitumor immunity by Ad-BD2-E1A. The results of this study indicate that in situ vaccination with the oncolytic BD2-expressing adenovirus preferentially attracts pDCs and promotes their maturation, and thus elicits potent tumor-specific immunity. This vaccine represents an attractive therapeutic strategy for the induction of individualized antitumor immunity.
Figures
Similar articles
-
Targeting the intratumoral dendritic cells by the oncolytic adenoviral vaccine expressing RANTES elicits potent antitumor immunity.J Immunother. 2009 Feb-Mar;32(2):145-56. doi: 10.1097/CJI.0b013e318193d31e. J Immunother. 2009. PMID: 19238013 Free PMC article.
-
The antitumor efficacy of IL-24 mediated by E1A and E1B triple regulated oncolytic adenovirus.Cancer Biol Ther. 2010 Aug 1;10(3):242-50. doi: 10.4161/cbt.10.3.12308. Epub 2010 Aug 10. Cancer Biol Ther. 2010. PMID: 20574150
-
E1A- and E1B-Double mutant replicating adenovirus elicits enhanced oncolytic and antitumor effects.Hum Gene Ther. 2007 Sep;18(9):773-86. doi: 10.1089/hum.2006.167. Hum Gene Ther. 2007. PMID: 17725410
-
Vaccination of fiber-modified adenovirus-transfected dendritic cells to express HER-2/neu stimulates efficient HER-2/neu-specific humoral and CTL responses and reduces breast carcinogenesis in transgenic mice.Cancer Gene Ther. 2008 Oct;15(10):655-66. doi: 10.1038/cgt.2008.18. Epub 2008 Apr 18. Cancer Gene Ther. 2008. PMID: 18421311
-
Virology- and immunology-based gene therapy for cancer.Cancer Immunol Immunother. 2006 Nov;55(11):1420-5. doi: 10.1007/s00262-006-0173-3. Epub 2006 May 12. Cancer Immunol Immunother. 2006. PMID: 16691360 Free PMC article. Review.
Cited by
-
Recent advances in oncolytic virus design.Clin Transl Oncol. 2011 Apr;13(4):229-39. doi: 10.1007/s12094-011-0647-4. Clin Transl Oncol. 2011. PMID: 21493183 Review.
-
Beta-defensin 2 enhances immunogenicity and protection of an adenovirus-based H5N1 influenza vaccine at an early time.Virus Res. 2013 Dec 26;178(2):398-403. doi: 10.1016/j.virusres.2013.09.013. Epub 2013 Sep 17. Virus Res. 2013. PMID: 24051000 Free PMC article.
-
Convergent evolution of defensin sequence, structure and function.Cell Mol Life Sci. 2017 Feb;74(4):663-682. doi: 10.1007/s00018-016-2344-5. Epub 2016 Aug 24. Cell Mol Life Sci. 2017. PMID: 27557668 Free PMC article. Review.
-
A role for B cells in facilitating defense against an NK cell-sensitive lung metastatic tumor is revealed by stress.J Neuroimmunol. 2017 Dec 15;313:99-108. doi: 10.1016/j.jneuroim.2017.10.017. Epub 2017 Nov 3. J Neuroimmunol. 2017. PMID: 29153616 Free PMC article.
-
Beyond cancer cells: Targeting the tumor microenvironment with gene therapy and armed oncolytic virus.Mol Ther. 2021 May 5;29(5):1668-1682. doi: 10.1016/j.ymthe.2021.04.015. Epub 2021 Apr 19. Mol Ther. 2021. PMID: 33845199 Free PMC article. Review.
References
-
- Hanada K, Yewdell JW., and , Yang JC. Immune recognition of a human renal cancer antigen through post-translational protein splicing. Nature. 2004;427:252–256. - PubMed
-
- Vigneron N, Stroobant V, Chapiro J, Ooms A, Degiovanni G, Morel S, et al. An antigenic peptide produced by peptide splicing in the proteasome. Science. 2004;304:587–590. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
