

An atlas of chaperone-protein interactions in Saccharomyces cerevisiae: implications to protein folding pathways in the cell
- PMID: 19536198
- PMCID: PMC2710862
- DOI: 10.1038/msb.2009.26
An atlas of chaperone-protein interactions in Saccharomyces cerevisiae: implications to protein folding pathways in the cell
Abstract
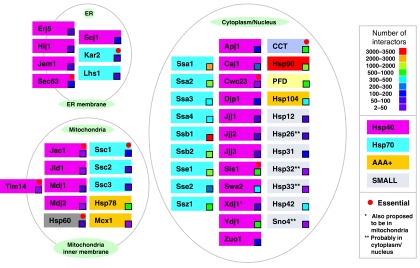
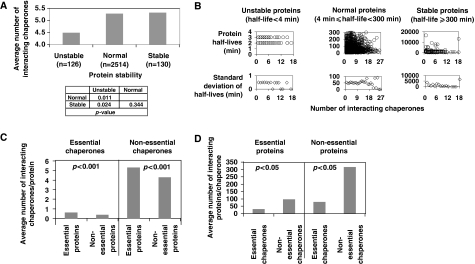
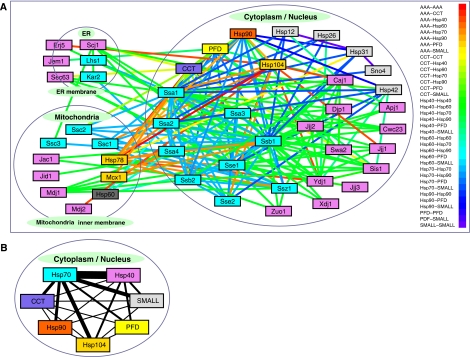
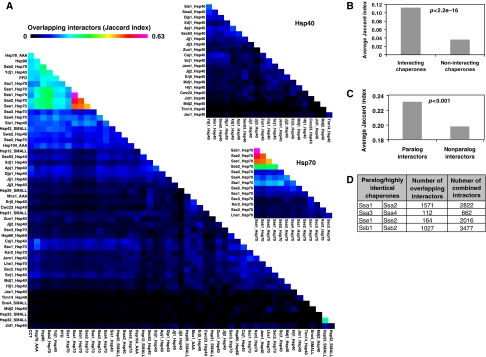
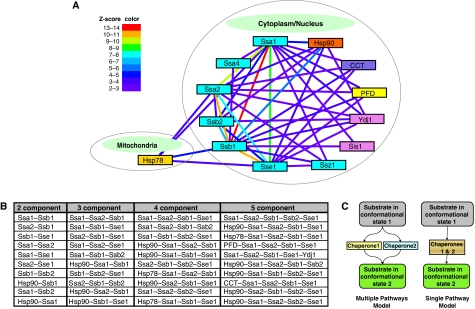
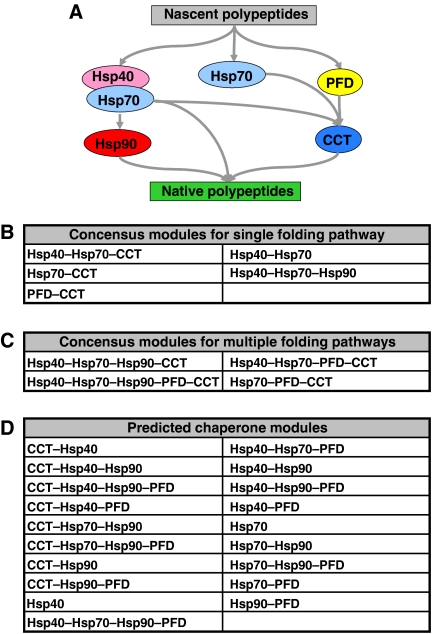
Molecular chaperones are known to be involved in many cellular functions, however, a detailed and comprehensive overview of the interactions between chaperones and their cofactors and substrates is still absent. Systematic analysis of physical TAP-tag based protein-protein interactions of all known 63 chaperones in Saccharomyces cerevisiae has been carried out. These chaperones include seven small heat-shock proteins, three members of the AAA+ family, eight members of the CCT/TRiC complex, six members of the prefoldin/GimC complex, 22 Hsp40s, 1 Hsp60, 14 Hsp70s, and 2 Hsp90s. Our analysis provides a clear distinction between chaperones that are functionally promiscuous and chaperones that are functionally specific. We found that a given protein can interact with up to 25 different chaperones during its lifetime in the cell. The number of interacting chaperones was found to increase with the average number of hydrophobic stretches of length between one and five in a given protein. Importantly, cellular hot spots of chaperone interactions are elucidated. Our data suggest the presence of endogenous multicomponent chaperone modules in the cell.
Conflict of interest statement
The authors declare that they have no conflict of interest.
Figures

References
-
- Albanese V, Yam AY, Baughman J, Parnot C, Frydman J (2006) Systems analyses reveal two chaperone networks with distinct functions in eukaryotic cells. Cell 124: 75–88 - PubMed
-
- Altschul SF, Gish W, Miller W, Myers EW, Lipman DJ (1990) Basic local alignment search tool. J Mol Biol 215: 403–410 - PubMed
-
- Ashburner M, Ball CA, Blake JA, Botstein D, Butler H, Cherry JM, Davis AP, Dolinski K, Dwight SS, Eppig JT, Harris MA, Hill DP, Issel-Tarver L, Kasarskis A, Lewis S, Matese JC, Richardson JE, Ringwald M, Rubin GM, Sherlock G (2000) Gene ontology: tool for the unification of biology. The Gene Ontology Consortium. Nat Genet 25: 25–29 - PMC - PubMed
-
- Bader GD, Hogue CW (2002) Analyzing yeast protein-protein interaction data obtained from different sources. Nat Biotechnol 20: 991–997 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
