

Gene targeting of a disease-related gene in human induced pluripotent stem and embryonic stem cells
- PMID: 19540188
- PMCID: PMC2720132
- DOI: 10.1016/j.stem.2009.05.023
Gene targeting of a disease-related gene in human induced pluripotent stem and embryonic stem cells
Abstract
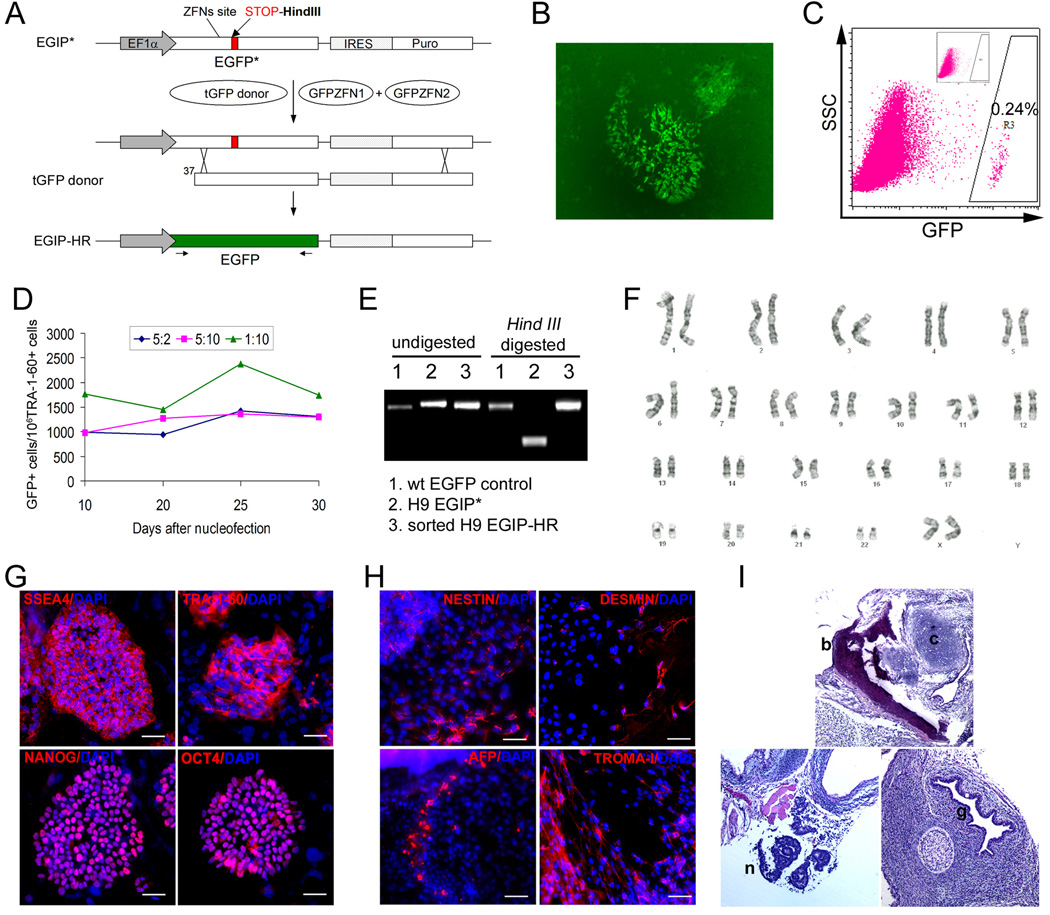
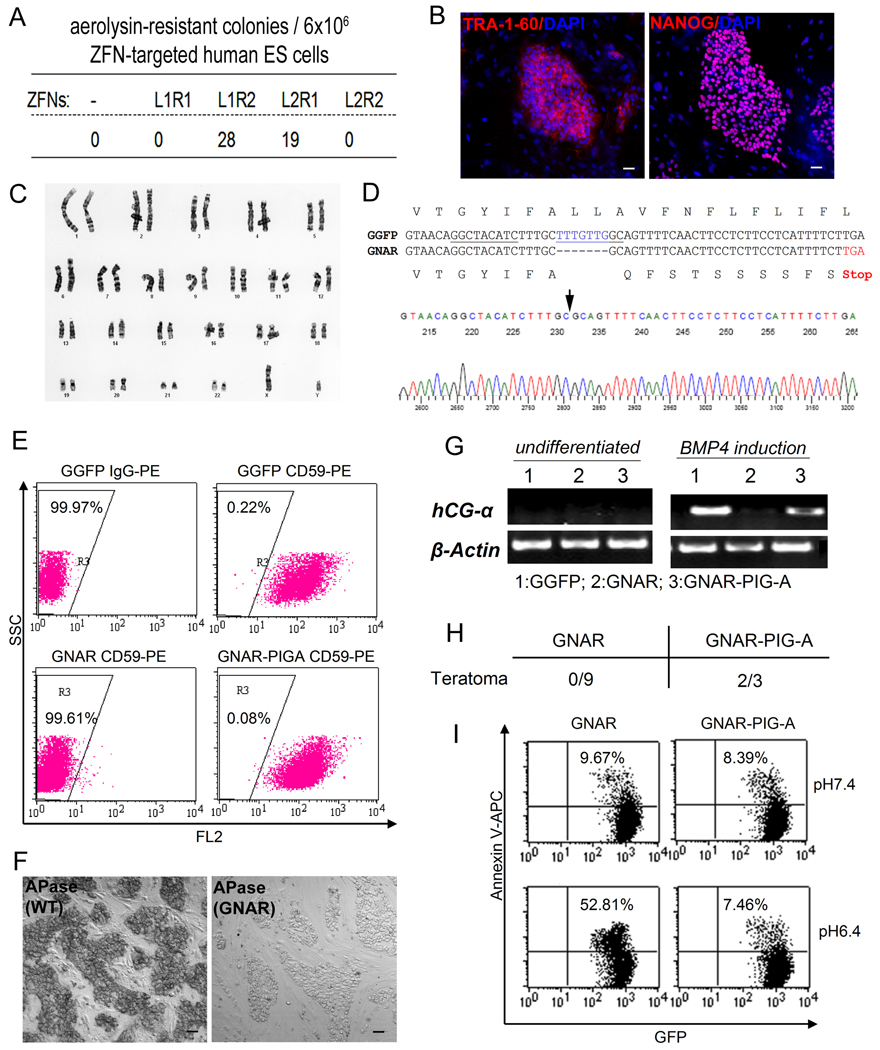
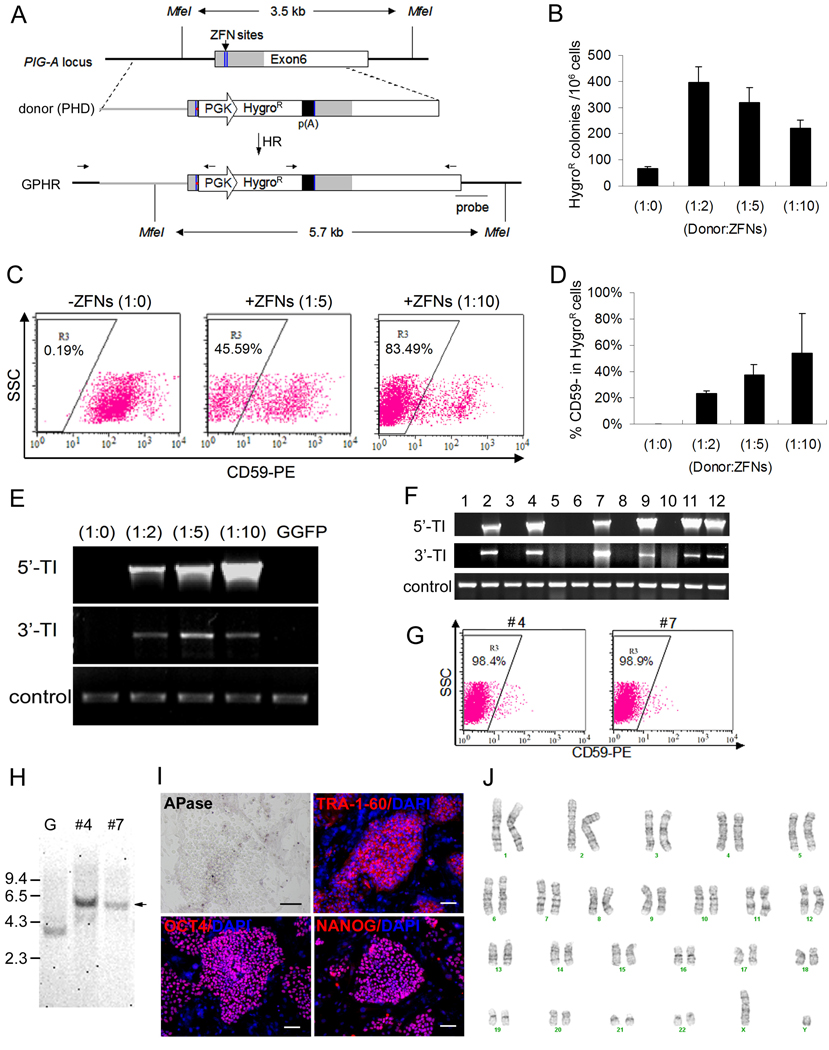
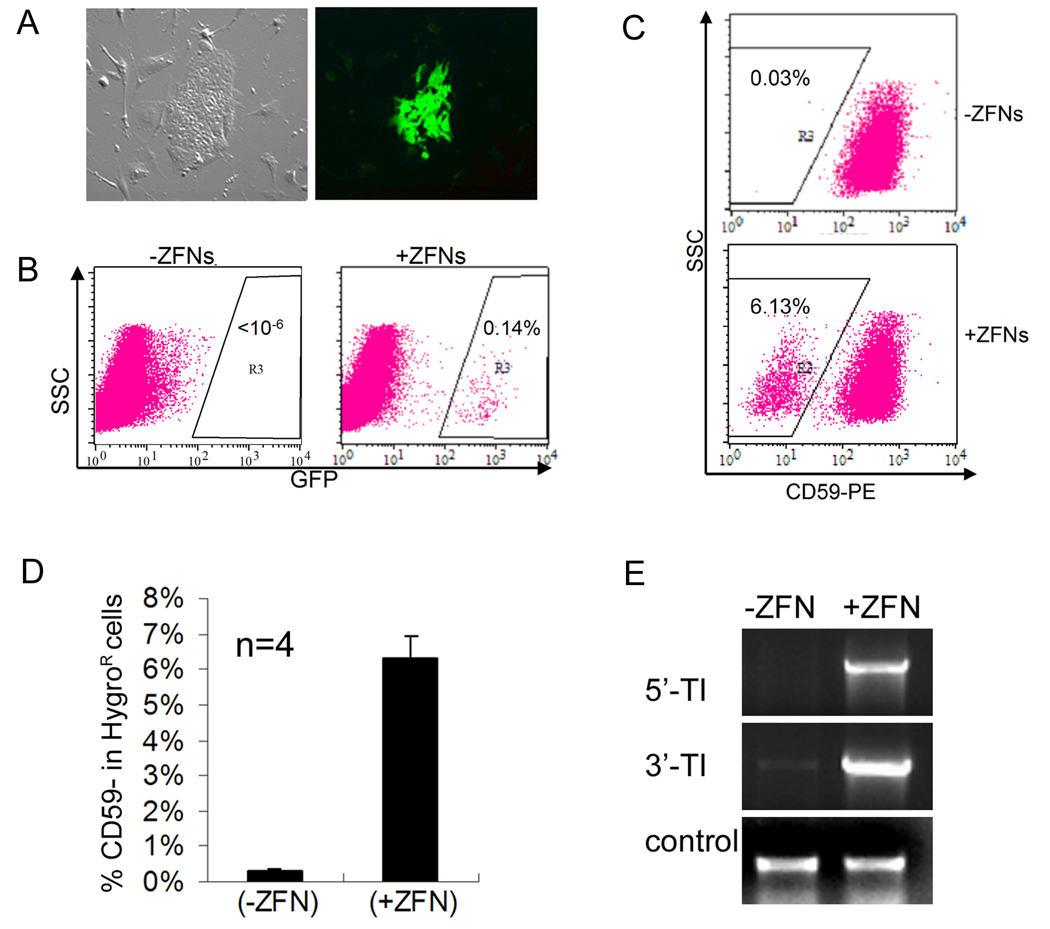
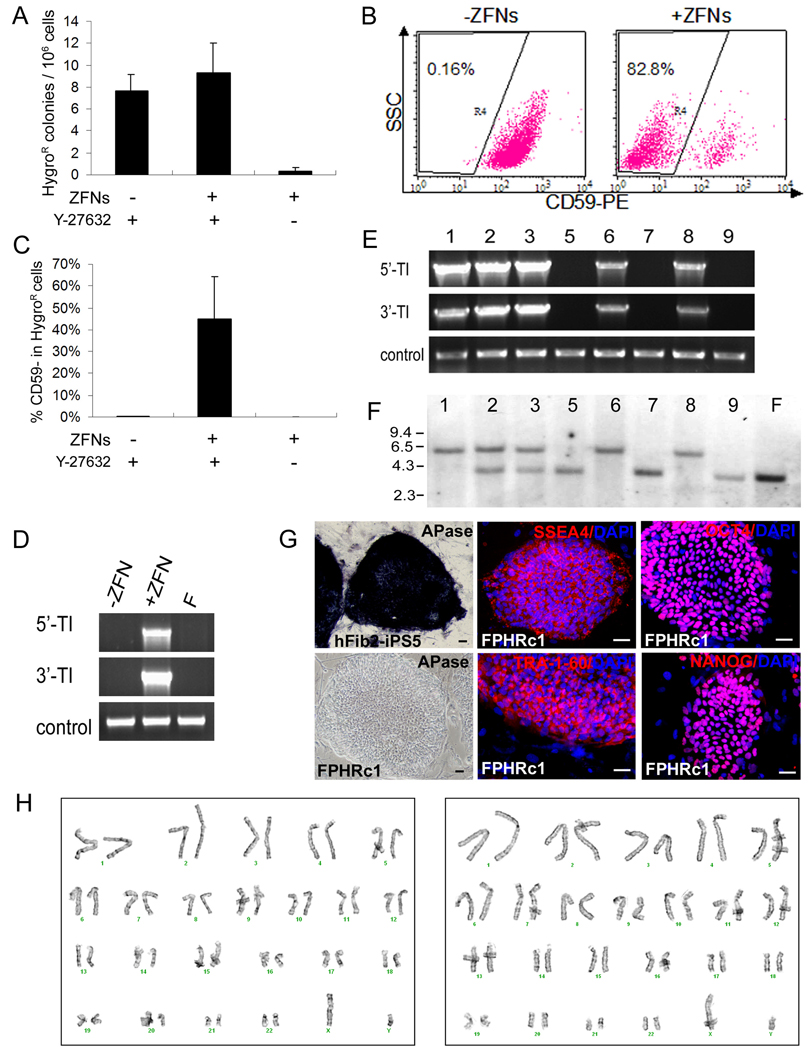
We report here homologous recombination (HR)-mediated gene targeting of two different genes in human iPS cells (hiPSCs) and human ES cells (hESCs). HR-mediated correction of a chromosomally integrated mutant GFP reporter gene reaches efficiencies of 0.14%-0.24% in both cell types transfected by donor DNA with plasmids expressing zinc finger nucleases (ZFNs). Engineered ZFNs that induce a sequence-specific double-strand break in the GFP gene enhanced HR-mediated correction by > 1400-fold without detectable alterations in stem cell karyotypes or pluripotency. Efficient HR-mediated insertional mutagenesis was also achieved at the endogenous PIG-A locus, with a > 200-fold enhancement by ZFNs targeted to that gene. Clonal PIG-A null hESCs and iPSCs with normal karyotypes were readily obtained. The phenotypic and biological defects were rescued by PIG-A transgene expression. Our study provides the first demonstration of HR-mediated gene targeting in hiPSCs and shows the power of ZFNs for inducing specific genetic modifications in hiPSCs, as well as hESCs.
Figures
References
-
- Cai L, Ye Z, Zhou BY, Mali P, Zhou C, Cheng L. Promoting human embryonic stem cell renewal or differentiation by modulating Wnt signal and culture conditions. Cell Res. 2007;17:62–72. - PubMed
-
- Cathomen T, Joung KJ. Zinc-finger nucleases: the next generation emerges. Mol Ther. 2008;16:1200–1207. - PubMed
-
- Costa M, Dottori M, Sourris K, Jamshidi P, Hatzistavrou T, Davis R, Azzola L, Jackson S, Lim SM, Pera M, et al. A method for genetic modification of human embryonic stem cells using electroporation. Nat Protoc. 2007;2:792–796. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
