

Multiple pontomedullary mechanisms of respiratory rhythmogenesis
- PMID: 19540366
- PMCID: PMC2734878
- DOI: 10.1016/j.resp.2009.06.011
Multiple pontomedullary mechanisms of respiratory rhythmogenesis
Abstract
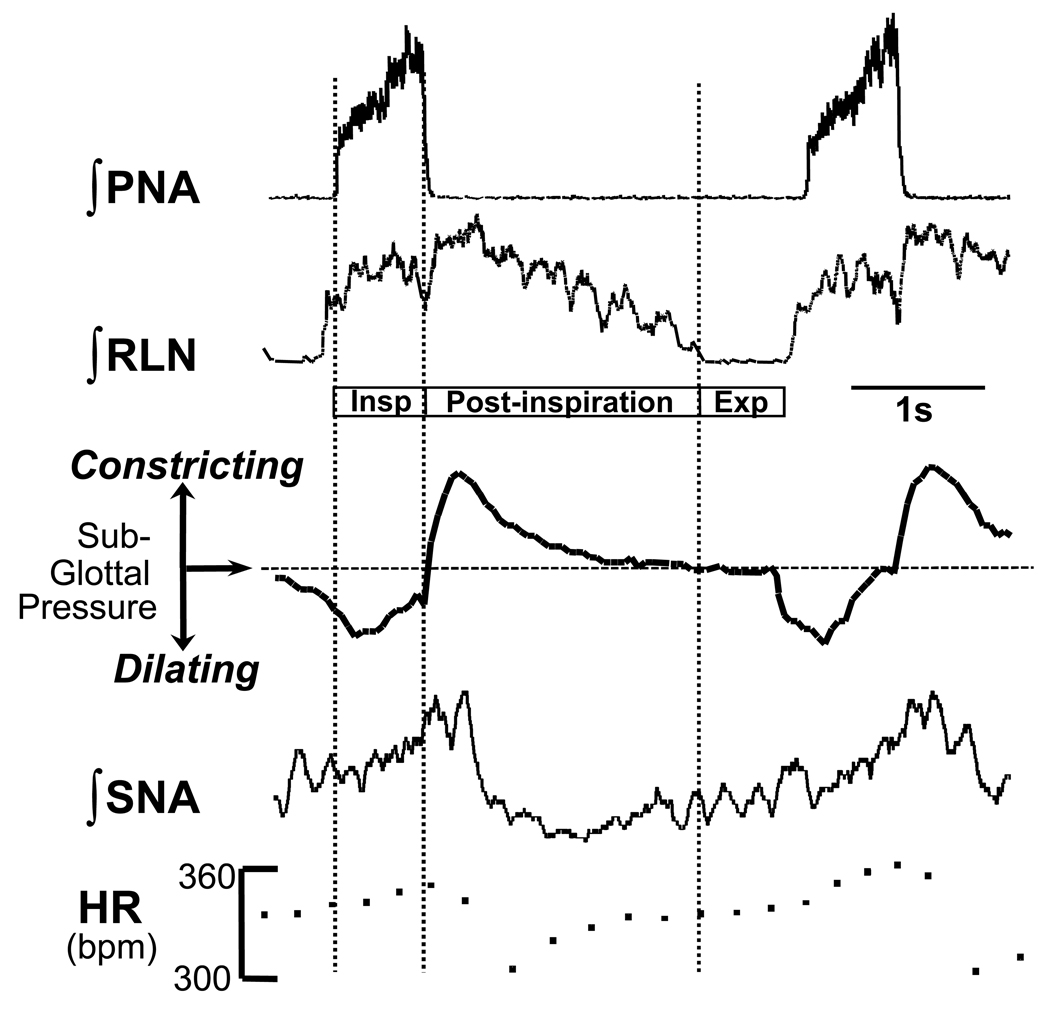
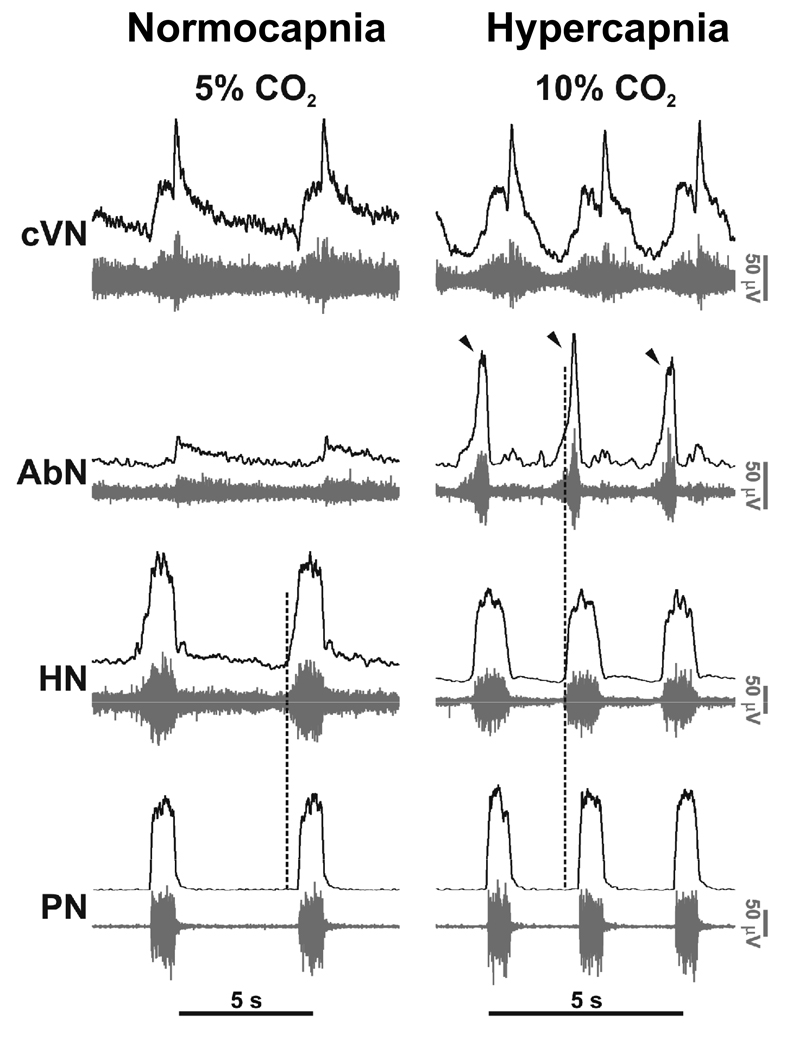
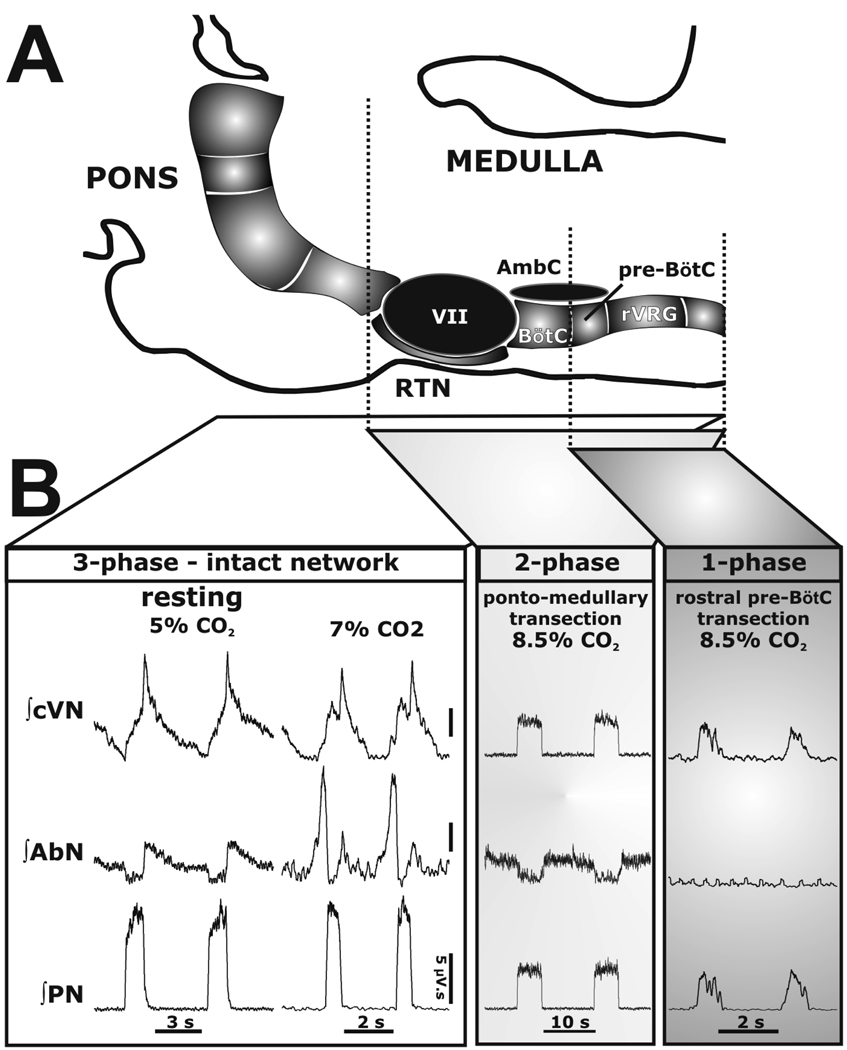
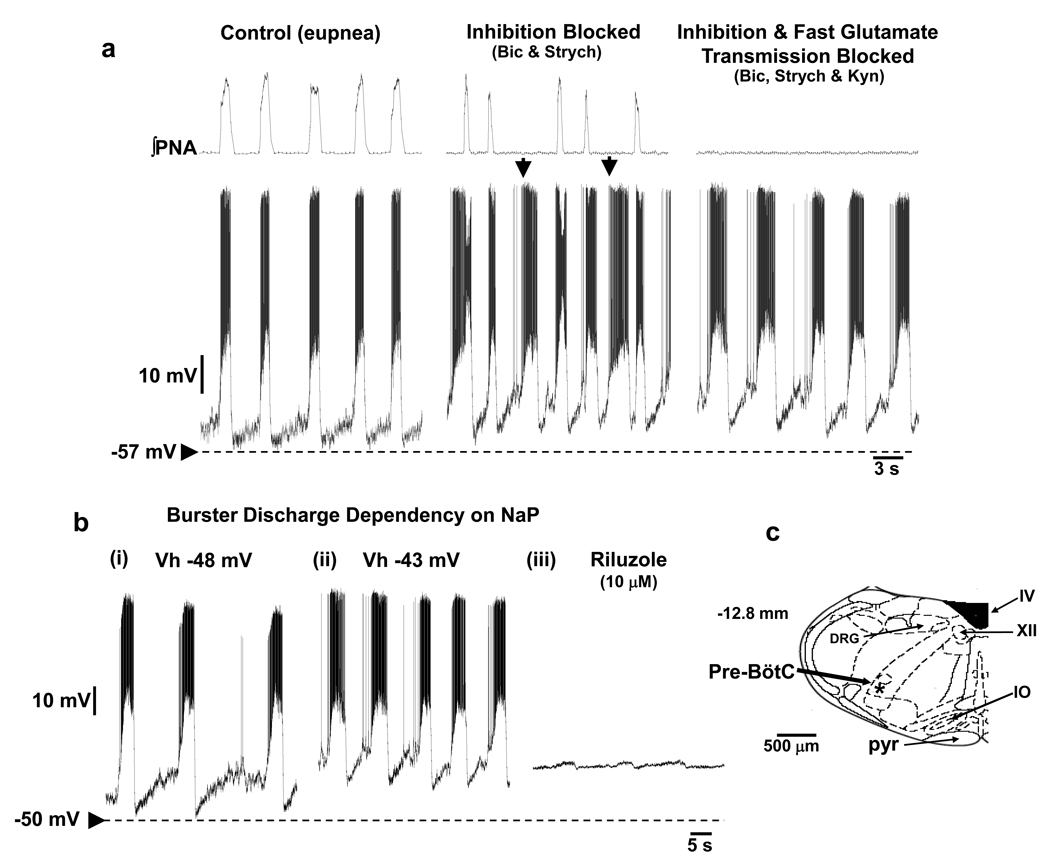
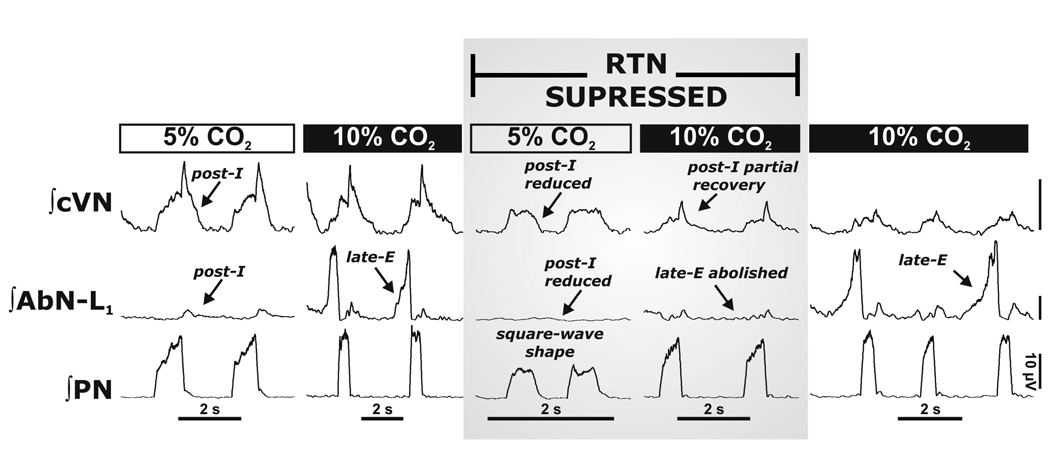
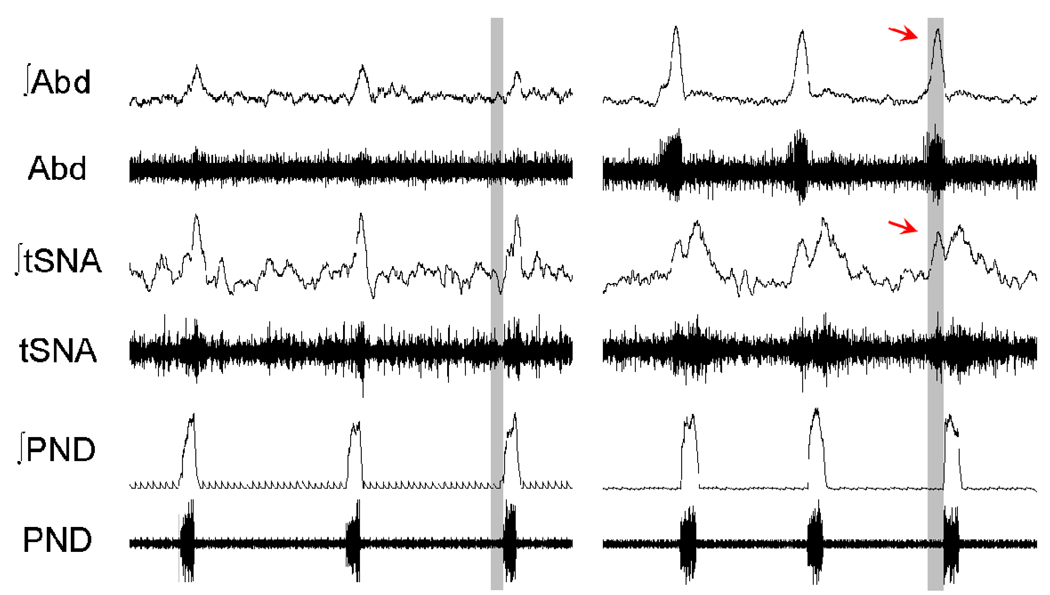
Mammalian central pattern generators producing rhythmic movements exhibit robust but flexible behavior. However, brainstem network architectures that enable these features are not well understood. Using precise sequential transections through the pons to medulla, it was observed that there was compartmentalization of distinct rhythmogenic mechanisms in the ponto-medullary respiratory network, which has rostro-caudal organization. The eupneic 3-phase respiratory pattern was transformed to a 2-phase and then to a 1-phase pattern as the network was physically reduced. The pons, the retrotrapezoid nucleus and glycine mediated inhibition are all essential for expression of the 3-phase rhythm. The 2-phase rhythm depends on inhibitory interactions (reciprocal) between Bötzinger and pre-Bötzinger complexes, whereas the 1-phase-pattern is generated within the pre-Bötzinger complex and is reliant on the persistent sodium current. In conditions of forced expiration, the RTN region was found to be essential for the expression of abdominal late expiratory activity. However, it is unknown whether the RTN generates or simply relays this activity. Entrained with the central respiratory network is the sympathetic nervous system, which exhibits patterns of discharge coupled with the respiratory cycle (in terms of both gain and phase of coupling) and dysfunctions in this coupling appear to underpin pathological conditions. In conclusion, the respiratory network has rhythmogenic capabilities at multiple levels of network organization, allowing expression of motor patterns specific for various physiological and pathophysiological respiratory behaviors.
Figures
References
-
- Anrep GV, Pascual W, Rossler R. Respiratory variations of the heart rate. II. The central mechanism of the sinus arrhythmia and the inter-relationships between central and reflex mechanism. Proc. Royal Soc. Lond. 1936;119:218–230. (Series B)
-
- Bianchi AL, Gestreau C. The respiratory network: An overview of a half century research. Respir. Physiol. Neurobiol. 2009 ?, ? - ? (This issue) - PubMed
-
- Büsselberg D, Bischoff AM, Paton JFR, Richter DW. Loss of glycinergic inhibition reveals two modes of respiratory rhythm generation. Pflügers Arch. 2001;441:444–449. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
