

A novel molecular switch
- PMID: 19540845
- PMCID: PMC2778020
- DOI: 10.1016/j.jmb.2009.06.039
A novel molecular switch
Abstract
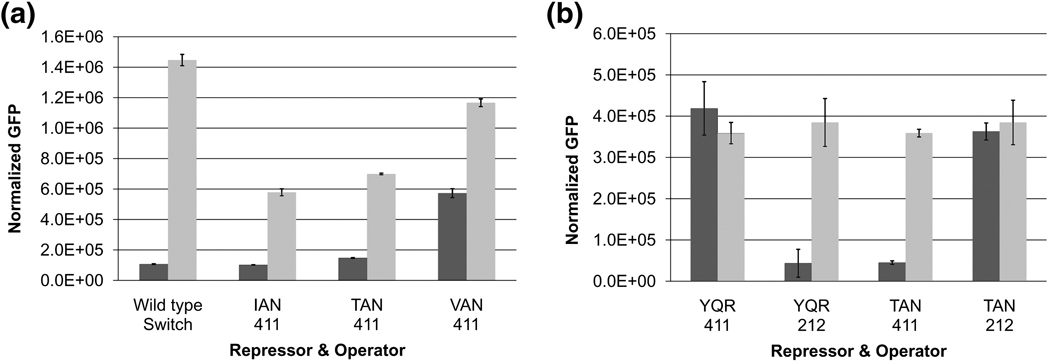
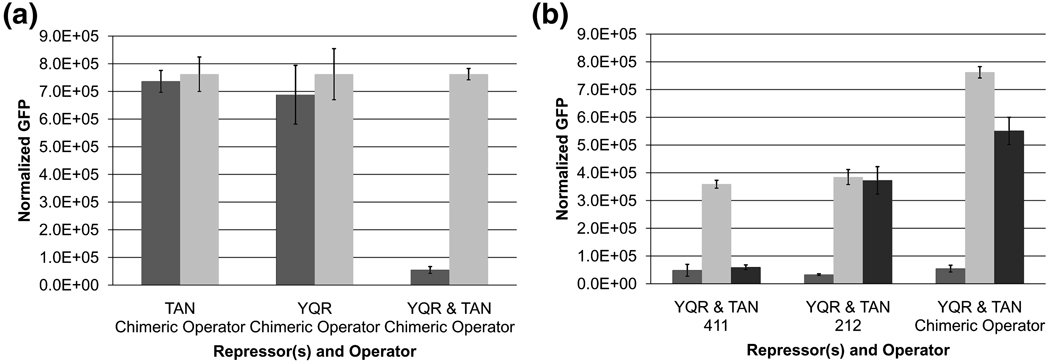
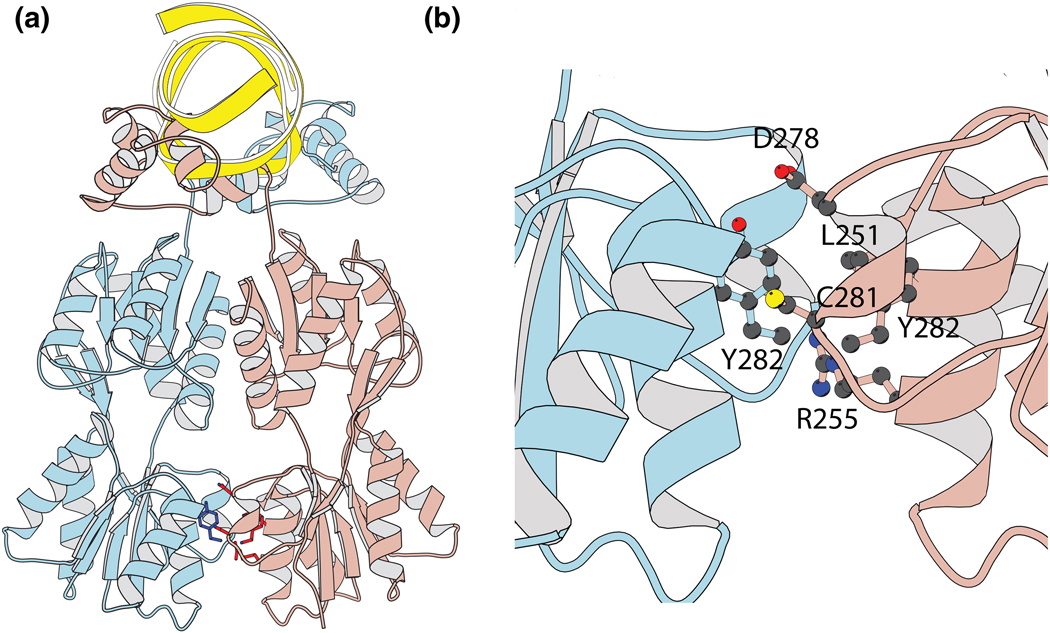
Transcriptional regulation is a fundamental process for regulating the flux of all metabolic pathways. For the last several decades, the lac operon has served as a valuable model for studying transcription. More recently, the switch that controls the operon has also been successfully adapted to function in mammalian cells. Here we describe how, using directed evolution, we have created a novel switch that recognizes an asymmetric operator sequence. The new switch has a repressor with altered headpiece domains for operator recognition and a redesigned dimer interface to create a heterodimeric repressor. Quite unexpectedly, the heterodimeric switch functions better than the natural system. It can repress more tightly than the naturally occurring switch of the lac operon; it is less leaky and can be induced more efficiently. Ultimately, these novel repressors could be evolved to recognize eukaryotic promoters and used to regulate gene expression in mammalian systems.
Figures



References
-
- Jacob F, Monod J. Genetic Regulatory Mechanisms in the Synthesis of Proteins. J. Mol. Biol. 1961;3:318–356. - PubMed
-
- Bell CE, Lewis M. The Lac repressor: a second generation of structural and functional studies. Current Opinion in Structural Biology. 2001;11:19–25. - PubMed
-
- Hu MC, Davidson N. The inducible lac operator-repressor system is functional in mammalian cells. Cell. 1987;48:555–566. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
