

Mechanism and evolution of DNA primases
- PMID: 19540940
- PMCID: PMC2846230
- DOI: 10.1016/j.bbapap.2009.06.011
Mechanism and evolution of DNA primases
Abstract
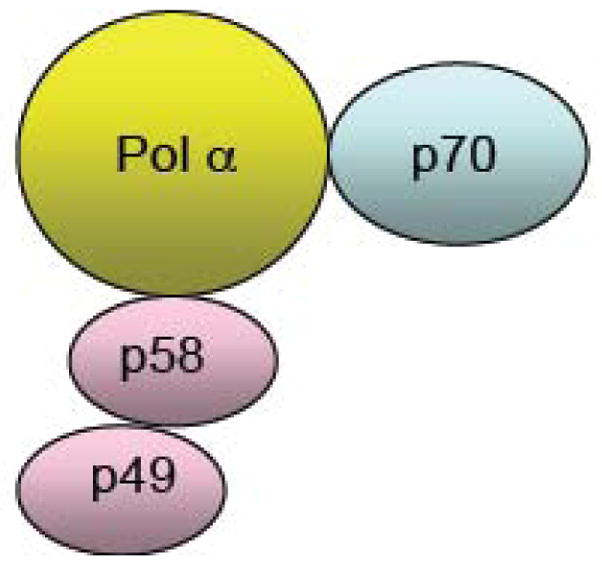
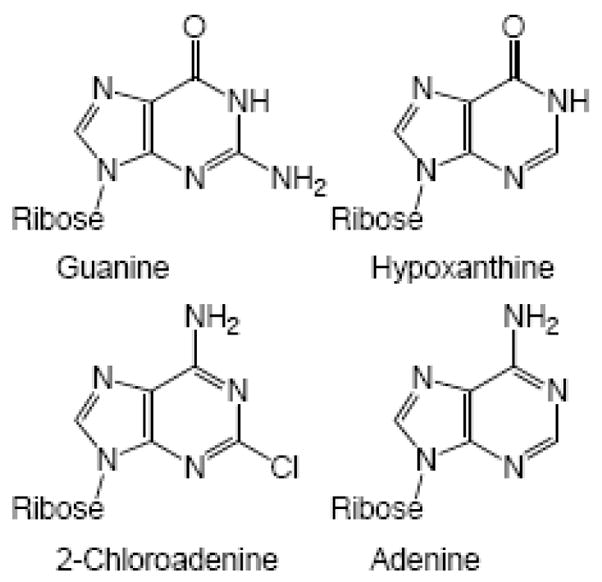
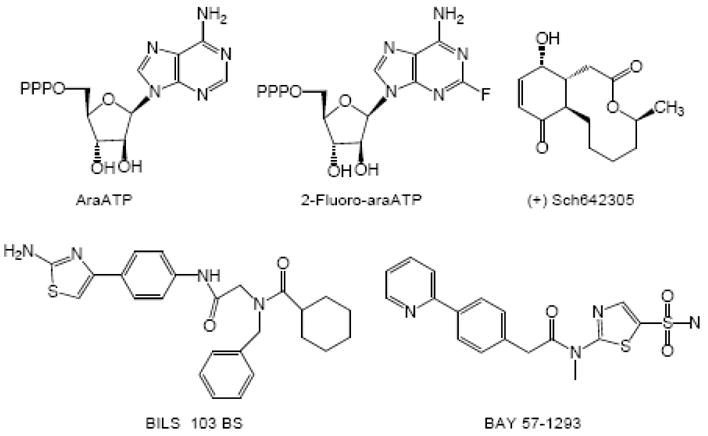
DNA primase synthesizes short RNA primers that replicative polymerases further elongate in order to initiate the synthesis of all new DNA strands. Thus, primase owes its existence to the inability of DNA polymerases to initiate DNA synthesis starting with 2 dNTPs. Here, we discuss the evolutionary relationships between the different families of primases (viral, eubacterial, archael, and eukaryotic) and the catalytic mechanisms of these enzymes. This includes how they choose an initiation site, elongate the growing primer, and then only synthesize primers of defined length via an inherent ability to count. Finally, the low fidelity of primases along with the development of primase inhibitors is described.
Copyright (c) 2010 Elsevier B.V. All rights reserved.
Figures






References
-
- Kornberg A, Baker T. DNA Replication. 2. W. H. Freeman & Co; San Francisco: 1992.
-
- Zhang SS, Grosse F. Accuracy of DNA Primase. Journal of Molecular Biology. 1990;216:475–479. - PubMed
-
- Hauschka PV. Analysis of Nucleotide Pools in Animal Cells. Methods in Cell Biology. 1973;3:362–462. - PubMed
-
- LeBreton M, Henneke G, Norais C, Flament D, Myllykallio H, Querellou J, Raffin JP. The Heterodimeric Primase from the Euryarchaeon Pyrococcus abyssi: a Multifunctional Enzyme for Initiation and Repair? J Mol Biol. 2007;374:1172–1185. - PubMed
-
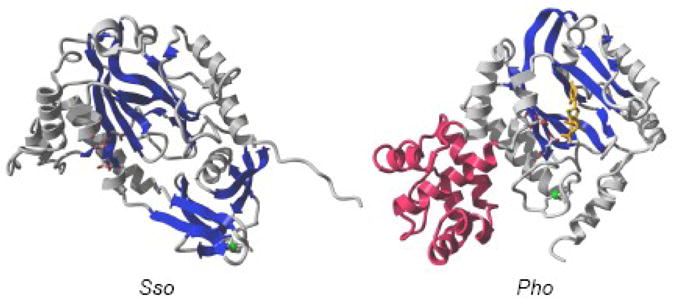
- Lao-Sireix SH, Bell SD. The Heterodimeric Primase of the Hyperthermophilic Archaeon Sulfolobus solfataricus Possesses DNA and RNA Primase, Polymerase, and 3′-terminal Nucleotidyl Transferase Activities. J Mol Biol. 2004;344:1251–1263. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
