

Semaphorin function in neural plasticity and disease
- PMID: 19541473
- PMCID: PMC2730419
- DOI: 10.1016/j.conb.2009.06.001
Semaphorin function in neural plasticity and disease
Abstract
The semaphorins, originally discovered as evolutionarily conserved steering molecules for developing axons, also influence neuronal structure and function in the early postnatal and juvenile nervous systems through several refinement processes. Semaphorins control synaptogenesis, axon pruning, and the density and maturation of dendritic spines. In addition, semaphorins and their downstream signaling components regulate synaptic physiology and neuronal excitability in the mature hippocampus, and these proteins are also implicated in a number of developmental, psychiatric, and neurodegenerative disorders. Significant inroads have been made in defining the mechanisms by which semaphorins regulate dynamic changes in the neuronal cytoskeleton at the molecular and cellular levels during embryonic nervous system development. However, comparatively little is known about how semaphorins influence neuronal structure and synaptic plasticity during adult nervous system homeostasis or following injury and disease. A detailed understanding of how semaphorins function beyond initial phases of neural network assembly is revealing novel insights into key aspects of nervous system physiology and pathology.
Conflict of interest statement
The authors declare no conflict of interest (RJP and RJG)
Figures
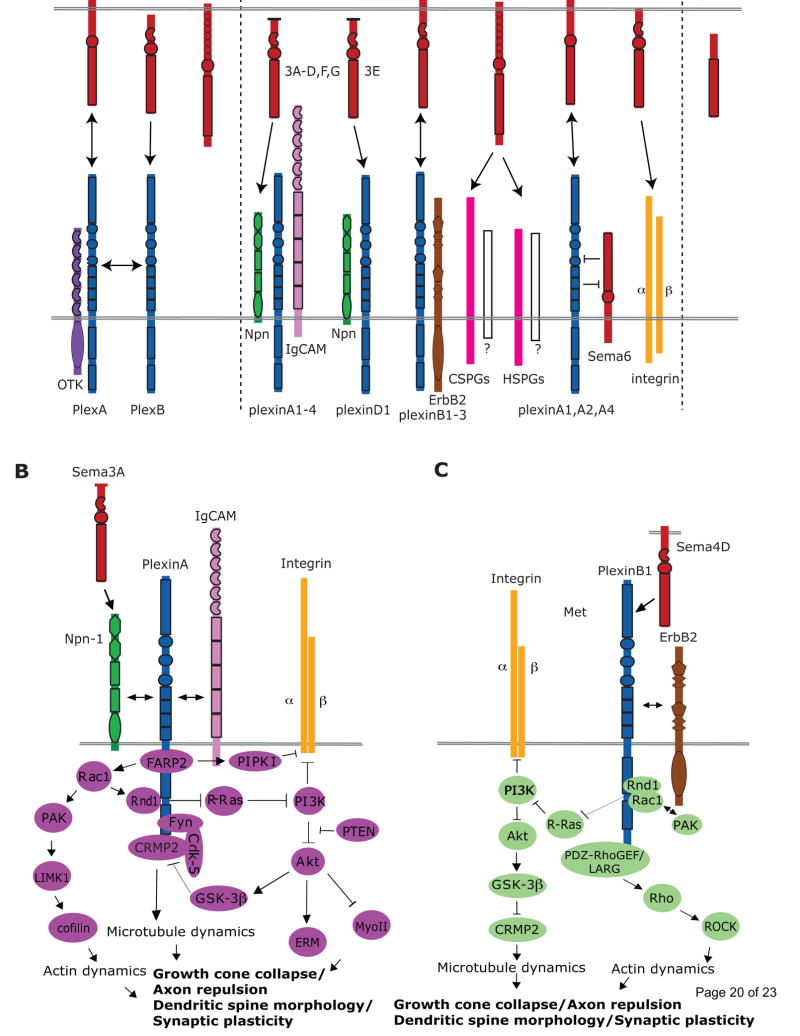
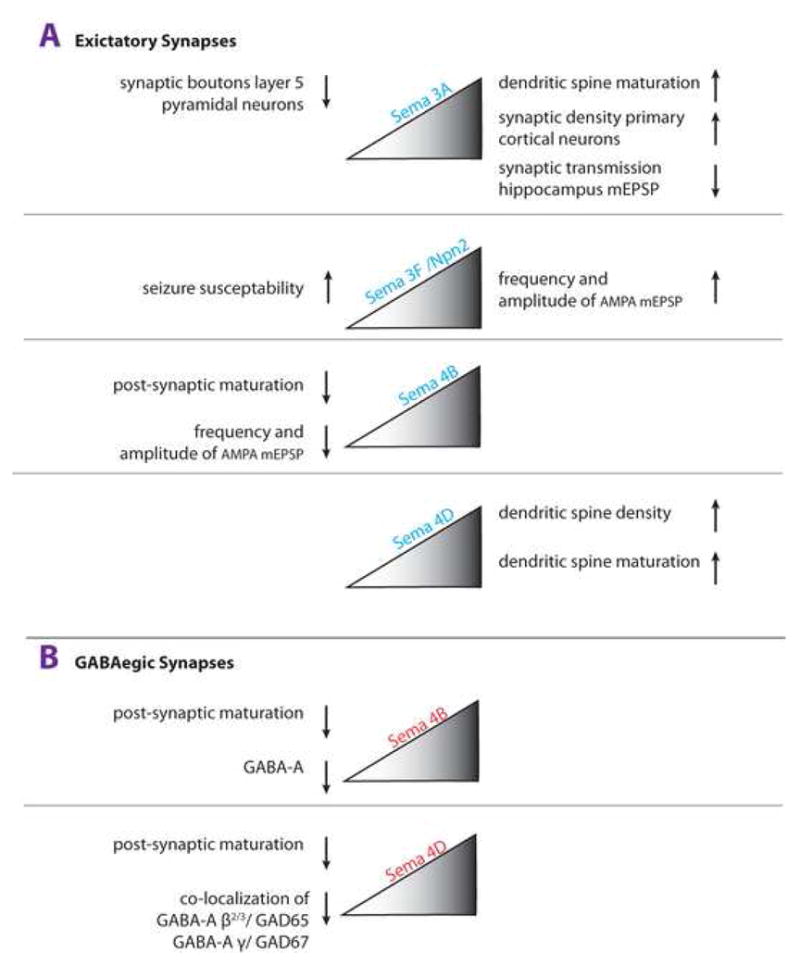
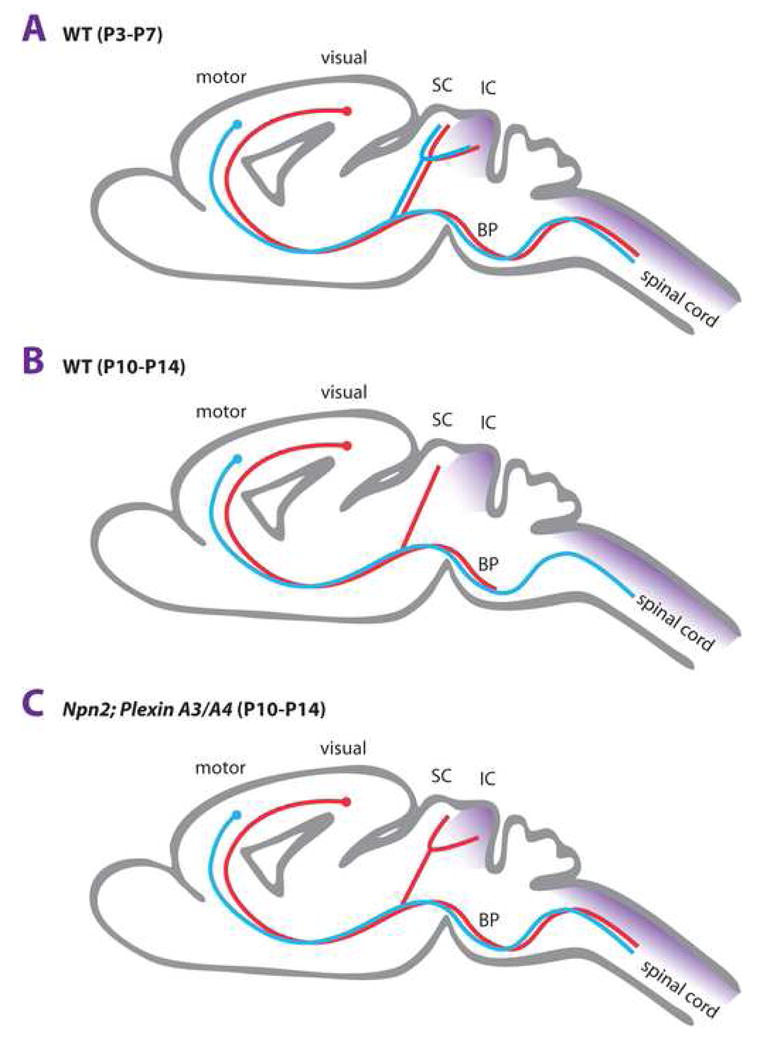
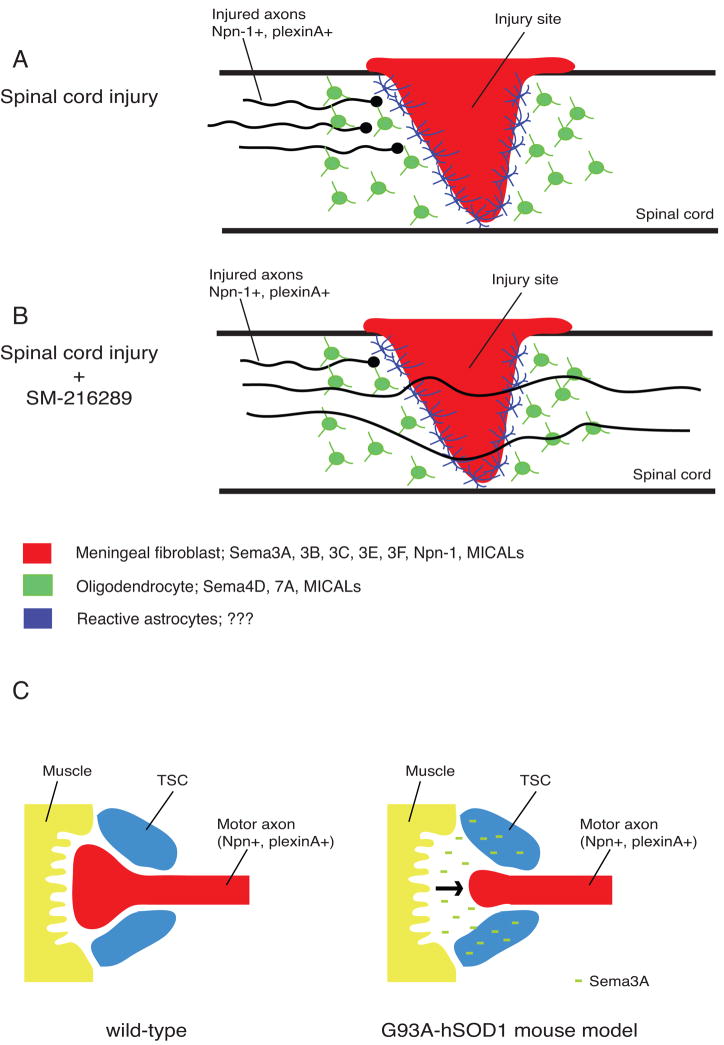
References
-
- Tran TS, Kolodkin AL, Bharadwaj R. Semaphorin regulation of cellular morphology. Annu Rev Cell Dev Biol. 2007;23:263–292. - PubMed
-
- Yuste R, Bonhoeffer T. Morphological changes in dendritic spines associated with long-term synaptic plasticity. Annu Rev Neurosci. 2001;24:1071–1089. - PubMed
-
- Falk J, Bechara A, Fiore R, Nawabi H, Zhou H, Hoyo-Becerra C, Bozon M, Rougon G, Grumet M, Puschel AW, et al. Dual functional activity of semaphorin 3B is required for positioning the anterior commissure. Neuron. 2005;48:63–75. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
