

CodY activates transcription of a small RNA in Bacillus subtilis
- PMID: 19542274
- PMCID: PMC2725614
- DOI: 10.1128/JB.00602-09
CodY activates transcription of a small RNA in Bacillus subtilis
Abstract
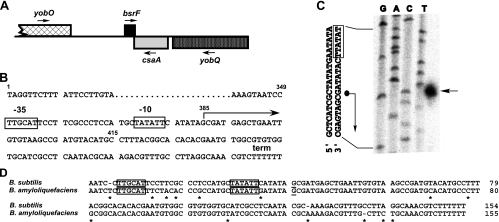
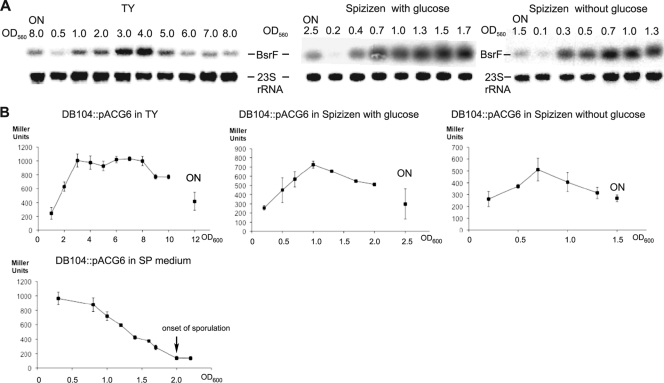
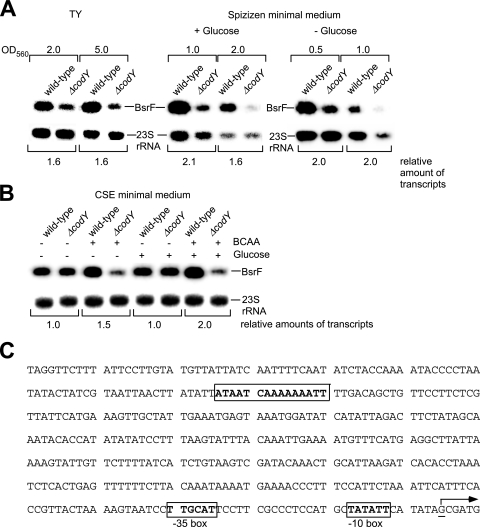
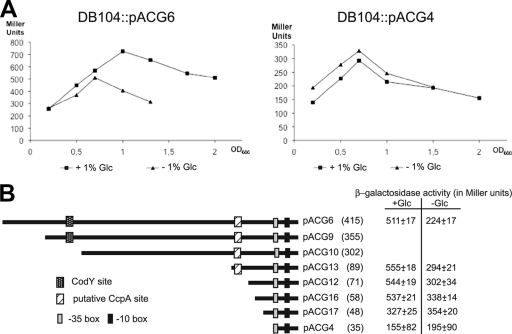
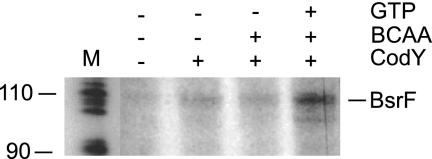
Regulatory small RNAs (sRNAs) in bacterial genomes have become a focus of research over the past 8 years. Whereas more than 100 such sRNAs have been found in Escherichia coli, relatively little is known about sRNAs in gram-positive bacteria. Using a computational approach, we identified two sRNAs in intergenic regions of the Bacillus subtilis genome, SR1 and SR2 (renamed BsrF). Recently, we demonstrated that SR1 inhibits the translation initiation of the transcriptional activator AhrC. Here, we describe detection of BsrF, its expression profile, and its regulation by CodY. Furthermore, we mapped the secondary structure of BsrF. BsrF is expressed in complex and minimal media in all growth phases in B. subtilis and, with a similar expression profile, also in Bacillus amyloliquefaciens. Neither overexpression nor deletion of bsrF affected the growth of B. subtilis. BsrF was found to be long-lived in complex and minimal media. Analysis of 13 putative transcription factor binding sites upstream of bsrF revealed only an effect for CodY. Here, we showed by using Northern blotting, lacZ reporter gene fusions, in vitro transcription, and DNase I footprinting that the transcription of bsrF is activated by CodY in the presence of branched-chain amino acids and GTP. Furthermore, BsrF transcription was increased 1.5- to 2-fold by glucose in the presence of branched-chain amino acids, and this increase was independent of the known glucose-dependent regulators. BsrF is the second target for which transcriptional activation by CodY has been discovered.
Figures




Similar articles
-
Activation of the Bacillus subtilis global regulator CodY by direct interaction with branched-chain amino acids.Mol Microbiol. 2004 Jul;53(2):599-611. doi: 10.1111/j.1365-2958.2004.04135.x. Mol Microbiol. 2004. PMID: 15228537
-
CodY is a nutritional repressor of flagellar gene expression in Bacillus subtilis.J Bacteriol. 2003 May;185(10):3118-26. doi: 10.1128/JB.185.10.3118-3126.2003. J Bacteriol. 2003. PMID: 12730172 Free PMC article.
-
Intracellular effectors regulating the activity of the Lactococcus lactis CodY pleiotropic transcription regulator.Mol Microbiol. 2004 Jul;53(2):613-21. doi: 10.1111/j.1365-2958.2004.04136.x. Mol Microbiol. 2004. PMID: 15228538
-
The CodY pleiotropic repressor controls virulence in gram-positive pathogens.FEMS Immunol Med Microbiol. 2011 Jul;62(2):123-39. doi: 10.1111/j.1574-695X.2011.00812.x. Epub 2011 May 27. FEMS Immunol Med Microbiol. 2011. PMID: 21539625 Review.
-
Intersection of the stringent response and the CodY regulon in low GC Gram-positive bacteria.Int J Med Microbiol. 2014 Mar;304(2):150-5. doi: 10.1016/j.ijmm.2013.11.013. Epub 2013 Dec 1. Int J Med Microbiol. 2014. PMID: 24462007 Review.
Cited by
-
The metabolic regulator CodY links Listeria monocytogenes metabolism to virulence by directly activating the virulence regulatory gene prfA.Mol Microbiol. 2015 Feb;95(4):624-44. doi: 10.1111/mmi.12890. Epub 2014 Dec 30. Mol Microbiol. 2015. PMID: 25430920 Free PMC article.
-
Hierarchical expression of genes controlled by the Bacillus subtilis global regulatory protein CodY.Proc Natl Acad Sci U S A. 2014 Jun 3;111(22):8227-32. doi: 10.1073/pnas.1321308111. Epub 2014 May 19. Proc Natl Acad Sci U S A. 2014. PMID: 24843172 Free PMC article.
-
Systems Level Analyses Reveal Multiple Regulatory Activities of CodY Controlling Metabolism, Motility and Virulence in Listeria monocytogenes.PLoS Genet. 2016 Feb 19;12(2):e1005870. doi: 10.1371/journal.pgen.1005870. eCollection 2016 Feb. PLoS Genet. 2016. PMID: 26895237 Free PMC article.
-
Integrative genomic analysis identifies isoleucine and CodY as regulators of Listeria monocytogenes virulence.PLoS Genet. 2012 Sep;8(9):e1002887. doi: 10.1371/journal.pgen.1002887. Epub 2012 Sep 6. PLoS Genet. 2012. PMID: 22969433 Free PMC article.
-
Optimization of Culture Conditions for Oxygen-Tolerant Regulatory [NiFe]-Hydrogenase Production from Ralstonia eutropha H16 in Escherichia coli.Microorganisms. 2021 May 31;9(6):1195. doi: 10.3390/microorganisms9061195. Microorganisms. 2021. PMID: 34073092 Free PMC article.
References
-
- Boisset, S., T. Geissmann, E. Huntzinger, P. Fechter, N. Bendridi, M. Possedko, C. Chevalier, A. C. Helfer, Y. Benito, A. Jacquier, C. Gaspin, F. Vandenesch, and P. Romby. 2007. Staphylococcus aureus RNAIII coordinately represses the synthesis of virulence factors and the transcription regulator Rot by an antisense mechanism. Genes Dev. 211353-1366. - PMC - PubMed
-
- Brantl, S. 1994. The copR gene product of plasmid pIP501 acts as a transcriptional repressor at the essential repR promotor. Mol. Microbiol. 14473-483. - PubMed
-
- Brantl, S. 2009. Bacterial chromosome-encoded small regulatory RNAs. Future Microbiol. 485-103. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
