

The transcriptional foundation of pluripotency
- PMID: 19542351
- PMCID: PMC2729344
- DOI: 10.1242/dev.024398
The transcriptional foundation of pluripotency
Abstract
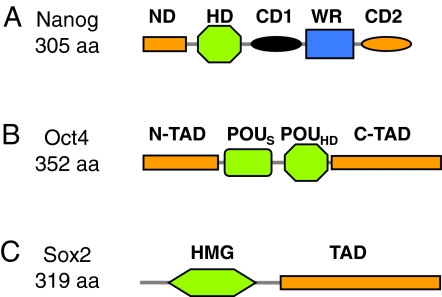
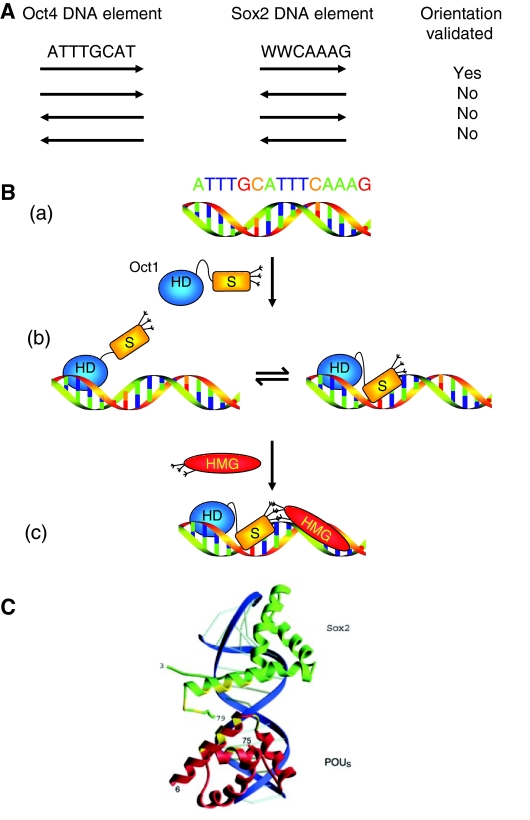
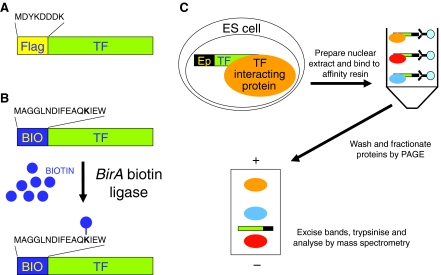
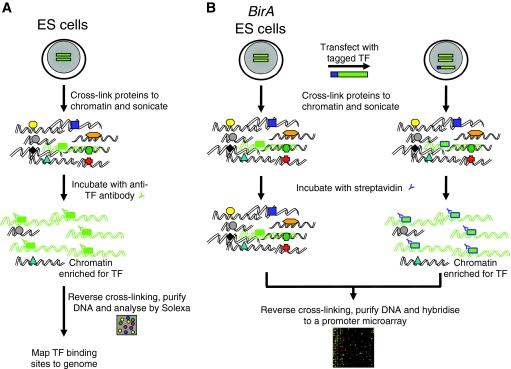
A fundamental goal in biology is to understand the molecular basis of cell identity. Pluripotent embryonic stem (ES) cell identity is governed by a set of transcription factors centred on the triumvirate of Oct4, Sox2 and Nanog. These proteins often bind to closely localised genomic sites. Recent studies have identified additional transcriptional modulators that bind to chromatin near sites occupied by Oct4, Sox2 and Nanog. This suggests that the combinatorial control of gene transcription might be fundamental to the ES cell state. Here we discuss how these observations advance our understanding of the transcription factor network that controls pluripotent identity and highlight unresolved issues that arise from these studies.
Figures
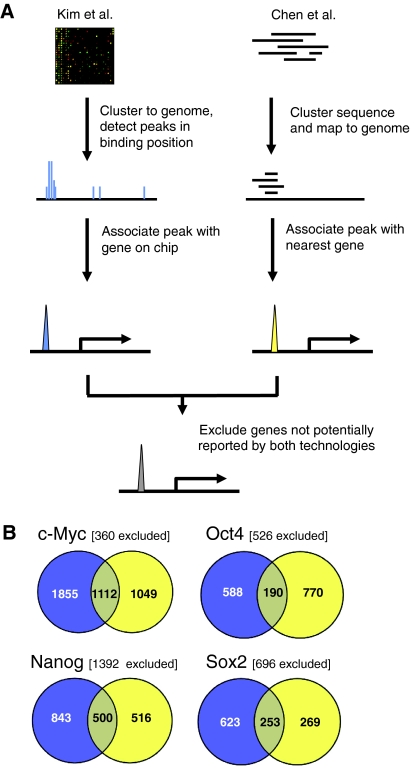
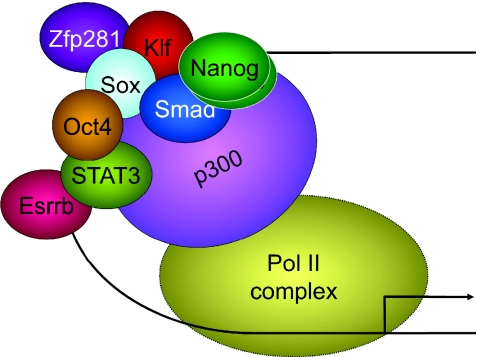
References
-
- Ambrosetti, D. C., Scholer, H. R., Dailey, L. and Basilico, C. (2000). Modulation of the activity of multiple transcriptional activation domains by the DNA binding domains mediates the synergistic action of Sox2 and Oct-3 on the fibroblast growth factor-4 enhancer. J. Biol. Chem. 275, 23387-23397. - PubMed
-
- Azuara, V., Perry, P., Sauer, S., Spivakov, M., Jorgensen, H. F., John, R. M., Gouti, M., Casanova, M., Warnes, G., Merkenschlager, M. et al. (2006). Chromatin signatures of pluripotent cell lines. Nat. Cell Biol. 8, 532-538. - PubMed
-
- Ben-Shushan, E., Thompson, J. R., Gudas, L. J. and Bergman, Y. (1998). Rex-1, a gene encoding a transcription factor expressed in the early embryo, is regulated via Oct-3/4 and Oct-6 binding to an octamer site and a novel protein, Rox-1, binding to an adjacent site. Mol. Cell. Biol. 18, 1866-1878. - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
