

IL-23 promotes the production of IL-17 by antigen-specific CD8 T cells in the absence of IL-12 and type-I interferons
- PMID: 19542449
- PMCID: PMC2729917
- DOI: 10.4049/jimmunol.0900939
IL-23 promotes the production of IL-17 by antigen-specific CD8 T cells in the absence of IL-12 and type-I interferons
Abstract
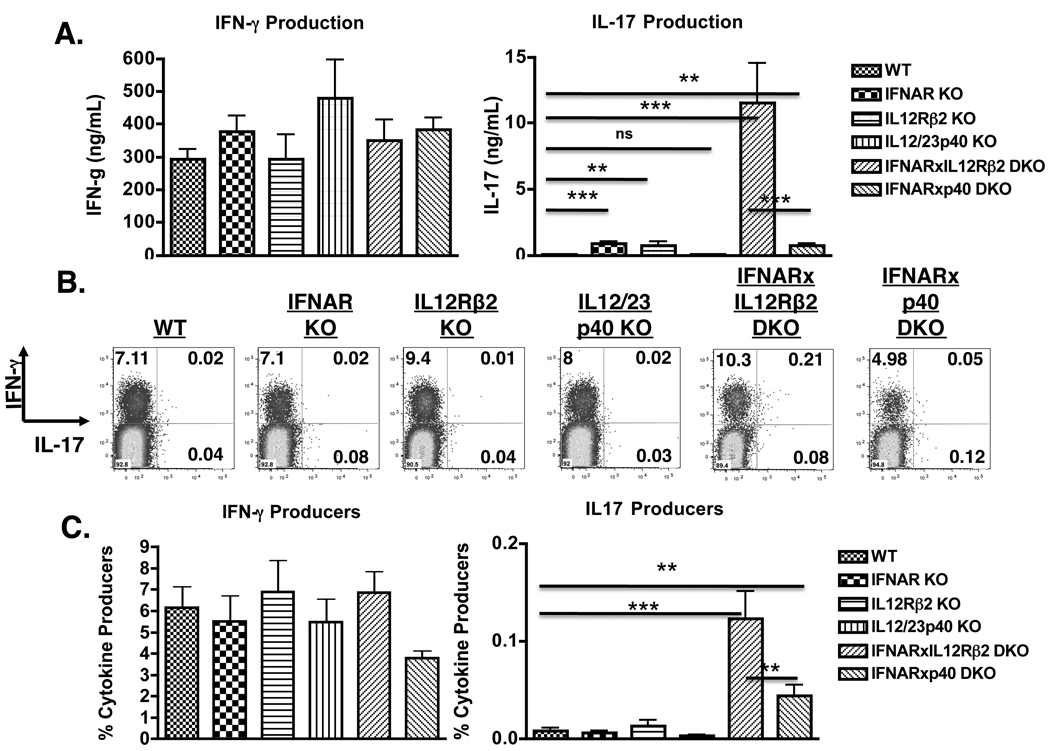
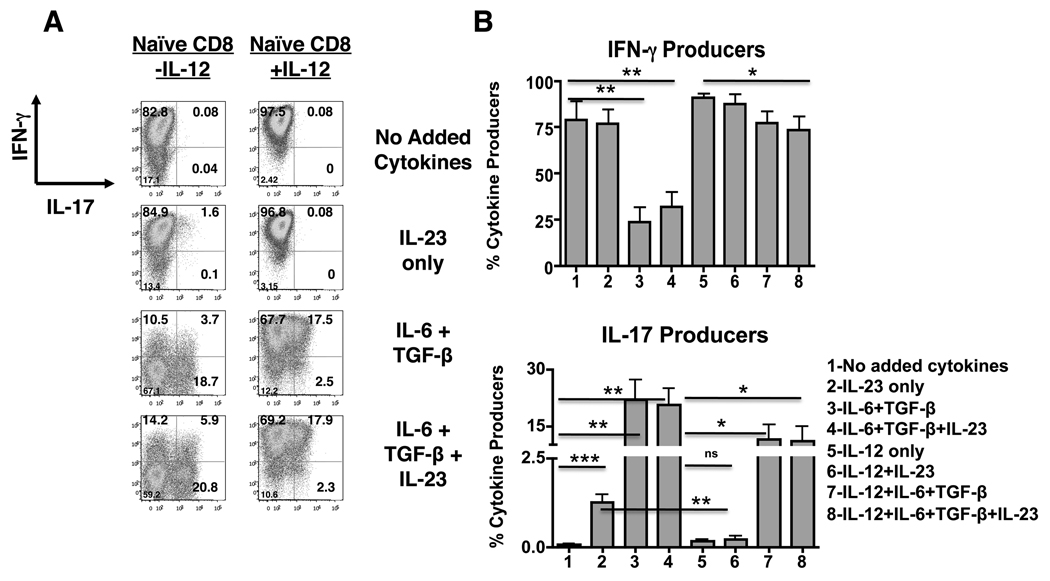
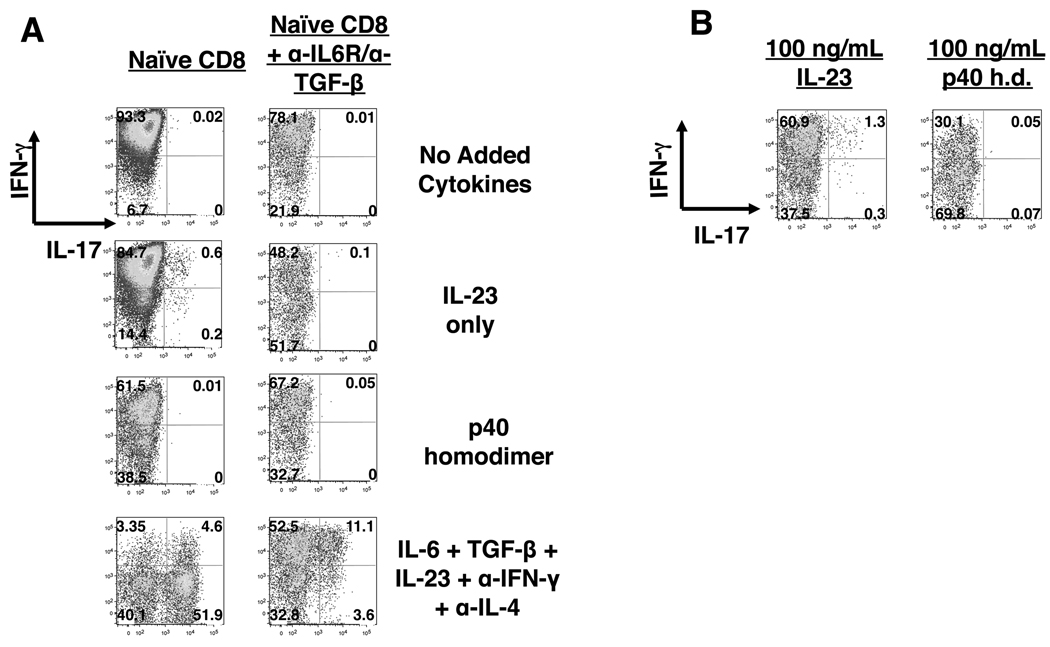
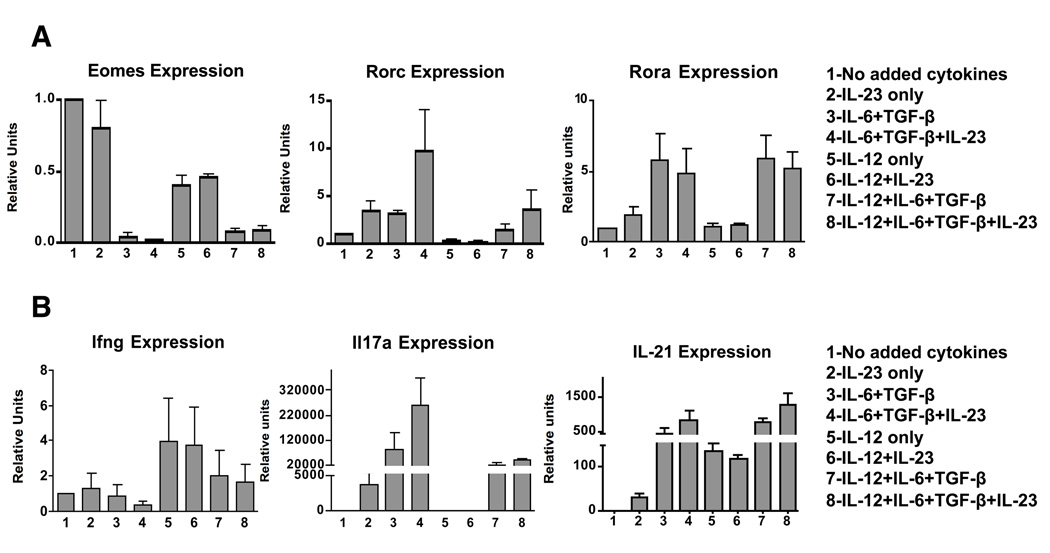
In contrast to CD4 T cells, CD8 T cells inherently differentiate into IFN-gamma-producing effectors. Accordingly, while generation of IFN-gamma-producing Th1 CD4 T cells was profoundly impaired in mice deficient for both type-I IFN and IL-12 signaling in response to infection with Listeria monocytogenes, generation of Ag-specific, IFN-gamma-producing CD8 T cells was unimpaired. However, a fraction of these CD8 T cells also produced IL-17 in an IL-23-dependent manner. Furthermore, the addition of IL-23 in vitro was sufficient for some naive CD8 T cells to differentiate into IFN-gamma/IL-17 dual-producing cells and was associated with increased expression of ROR-gammat and ROR-alpha. Addition of IL-6 and TGF-beta to IL-23 further augmented ROR-gammat and ROR-alpha expression and suppressed Eomes expression, thereby enhancing IL-17 production by CD8 T cells. A loss of cytotoxic function accompanied the production of IL-17, as the addition of IL-6 and TGF-beta resulted in a marked reduction of granzyme B and perforin expression. Thus, CD8 T cells retain sufficient plasticity to respond to environmental cues and can acquire additional effector functions in response to their environmental context.
Conflict of interest statement
The authors have no financial conflict of interest.
Figures



References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
