

IRAK4 kinase activity is required for Th17 differentiation and Th17-mediated disease
- PMID: 19542468
- PMCID: PMC3638260
- DOI: 10.4049/jimmunol.0802361
IRAK4 kinase activity is required for Th17 differentiation and Th17-mediated disease
Abstract
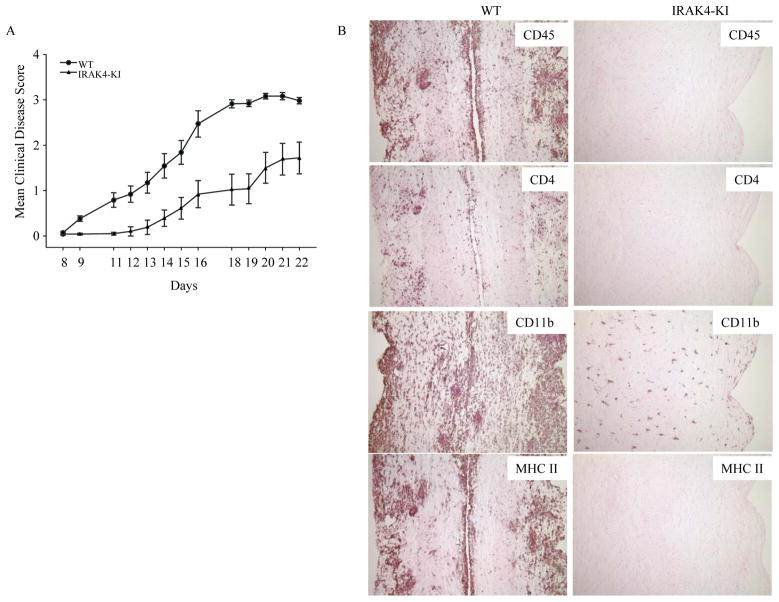
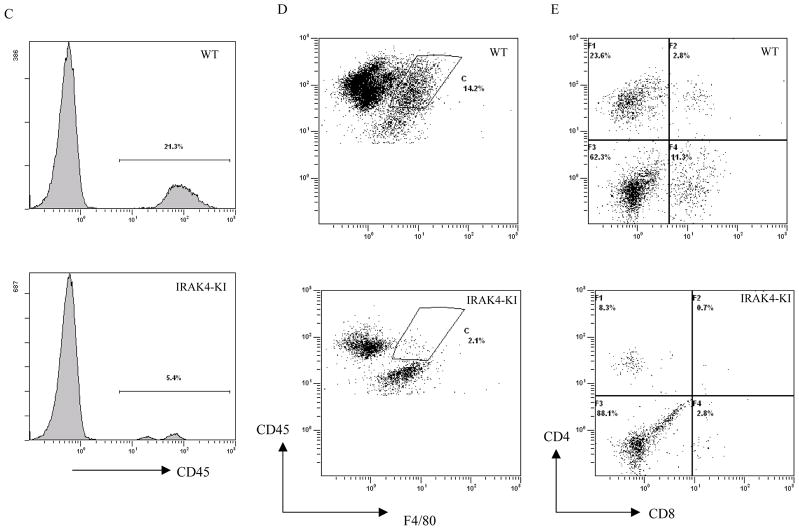
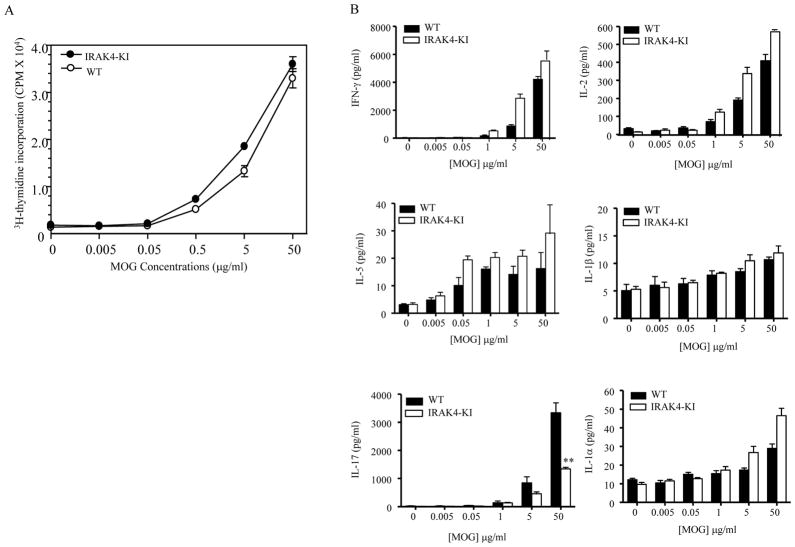
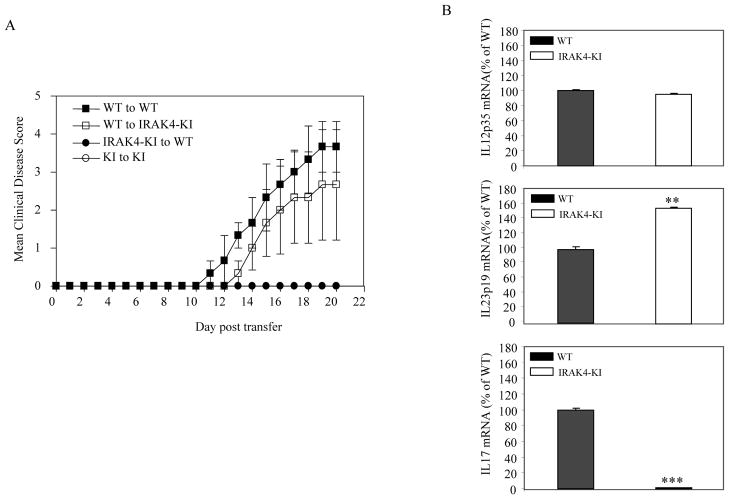
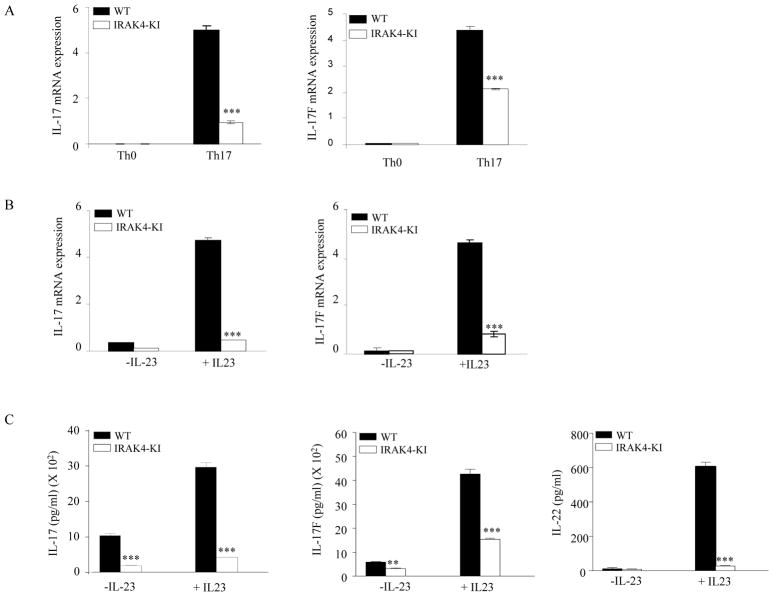
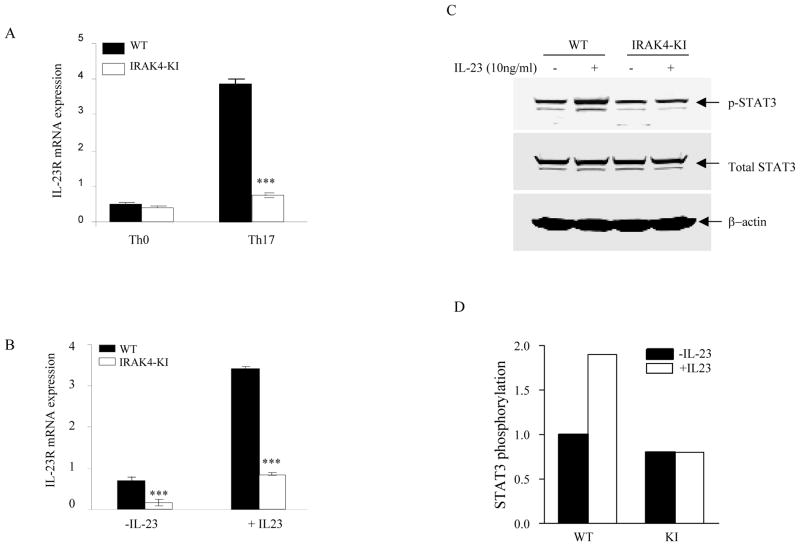
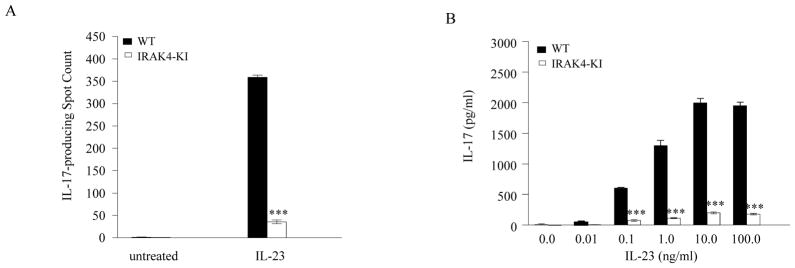
Both IL-23- and IL-1-mediated signaling pathways play important roles in Th17 cell differentiation, cytokine production, and autoimmune diseases. The IL-1R-associated kinase 4 (IRAK4) is critical for IL-1/TLR signaling. We show here that inactivation of IRAK4 kinase in mice (IRAK4 KI) results in significant resistance to experimental autoimmune encephalomyelitis due to a reduction in infiltrating inflammatory cells into the CNS and reduced Ag-specific CD4(+) T cell-mediated IL-17 production. Adoptive transfer of myelin oligodendrocyte glycoprotein 35-55-specific IRAK4 KI Th17 cells failed to induce experimental autoimmune encephalomyelitis in either wild-type or IRAK4 KI recipient mice, indicating the lack of autoantigen-specific Th17 cell activities in the absence of IRAK4 kinase activity. Furthermore, the absence of IRAK4 kinase activity blocked induction of IL-23R expression, STAT3 activation by IL-23, and Th17 cytokine expression in differentiated Th17 cells. Importantly, blockade of IL-1 signaling by IL-1RA inhibited Th17 differentiation and IL-23-induced cytokine expression in differentiated Th17 cells. The results of these studies demonstrate that IL-1-mediated IRAK4 kinase activity in T cells is essential for induction of IL-23R expression, Th17 differentiation, and autoimmune disease.
Figures
References
-
- Weaver CT, Hatton RD, Mangan PR, Harrington LE. IL-17 family cytokines and the expanding diversity of effector T cell lineages. Annu Rev Immunol. 2007;25:821–852. Review. - PubMed
-
- Weaver CT, Harrington LE, Mangan PR, Gavrieli M, Murphy KM. Th17: an effector CD4 T cell lineage with regulatory T cell ties. Immunity. 2006;24:677–688. Review. - PubMed
-
- Matusevicius D, Kivisäkk P, He B, Kostulas N, Ozenci V, Fredrikson S, Link H. Interleukin-17 mRNA expression in blood and CSF mononuclear cells is augmented in multiple sclerosis. Mult Scler. 1999;5:101–104. - PubMed
-
- Kim HR, Kim HS, Park MK, Cho ML, Lee SH, Kim HY. The clinical role of IL-23p19 in patients with rheumatoid arthritis. Scand J Rheumatol. 2007;36:259–264. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
