

Centromere assembly requires the direct recognition of CENP-A nucleosomes by CENP-N
- PMID: 19543270
- PMCID: PMC2704923
- DOI: 10.1038/ncb1899
Centromere assembly requires the direct recognition of CENP-A nucleosomes by CENP-N
Abstract
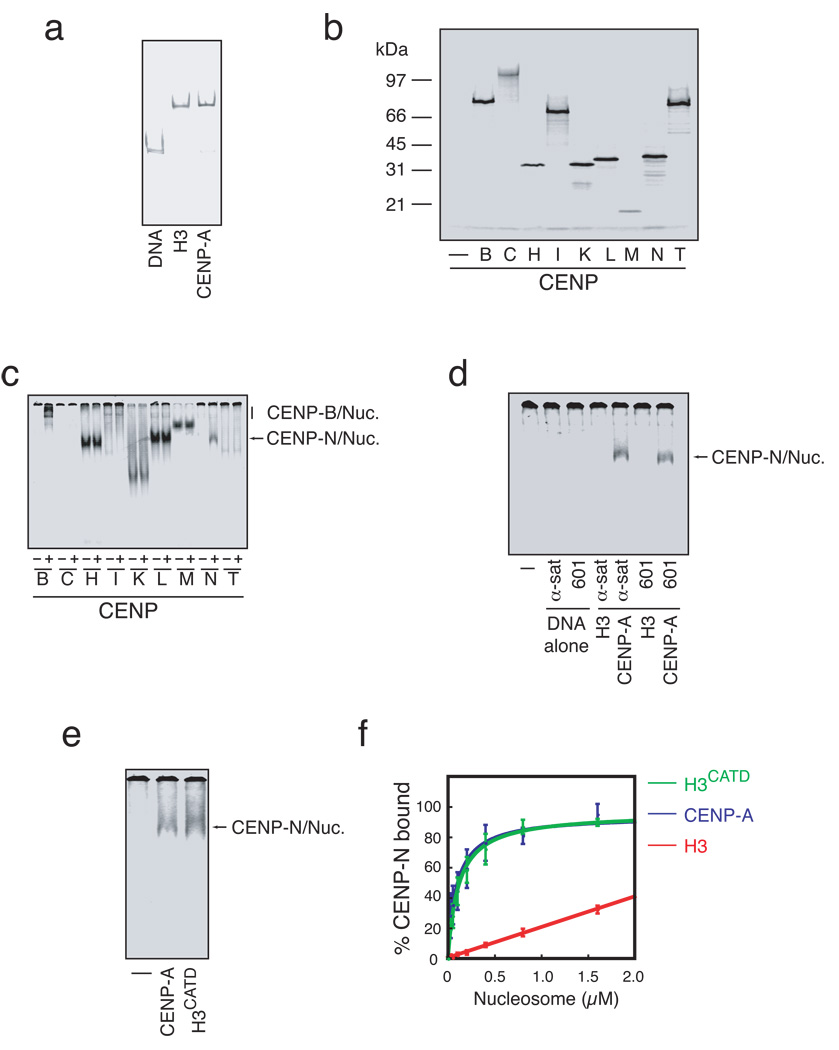
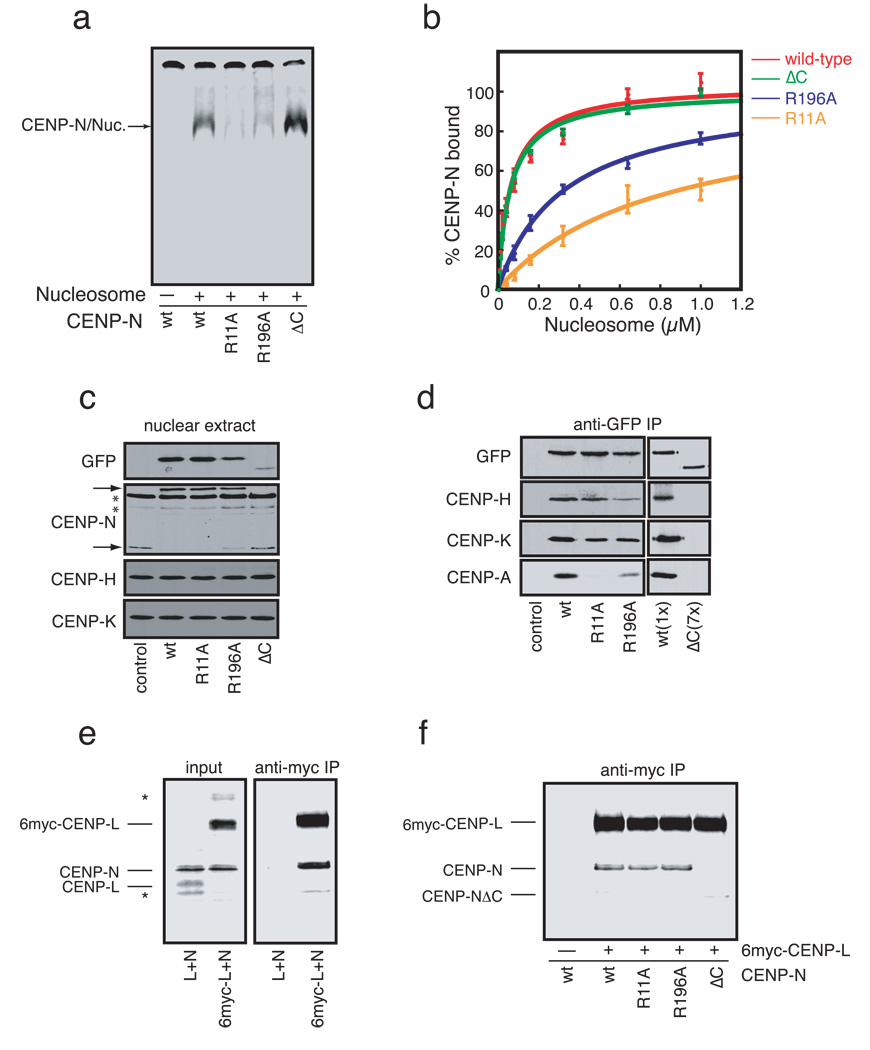
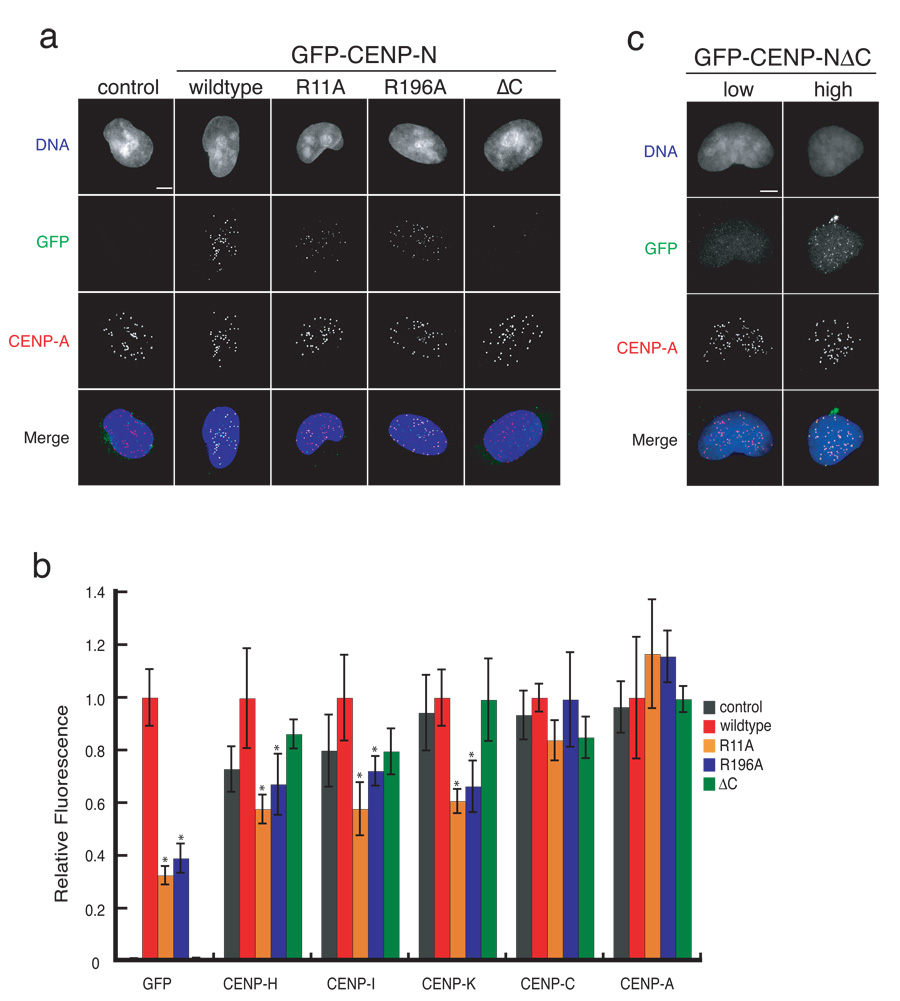
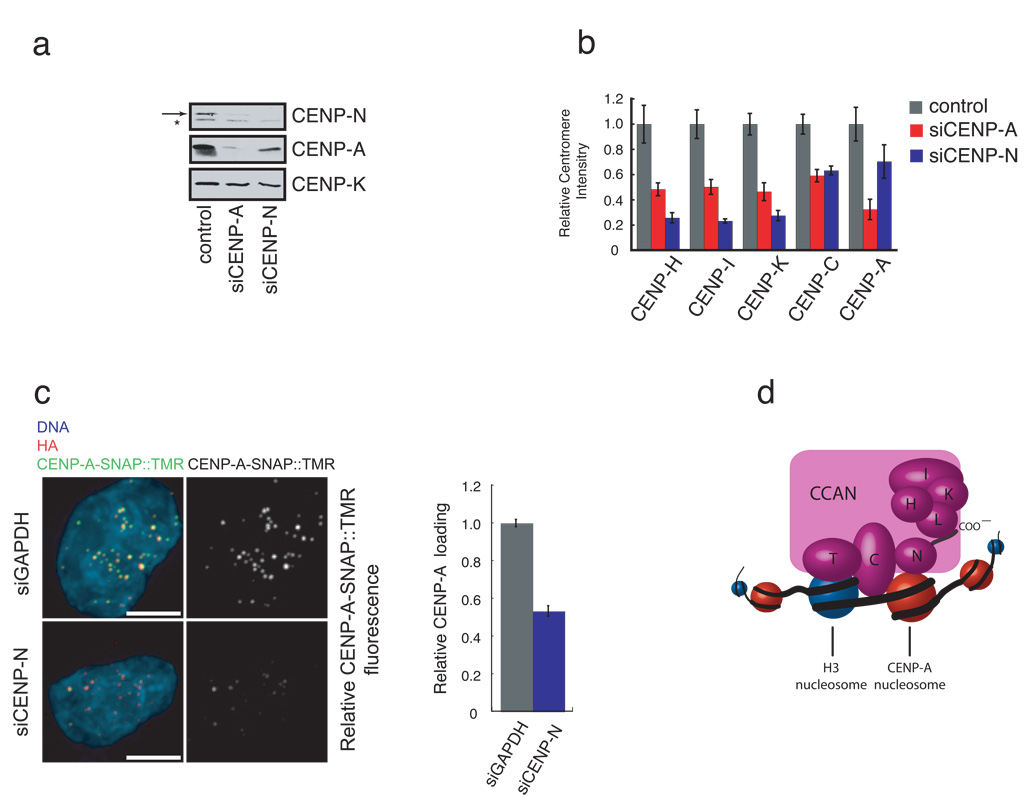
Centromeres are specialized chromosomal domains that direct kinetochore assembly during mitosis. CENP-A (centromere protein A), a histone H3-variant present exclusively in centromeric nucleosomes, is thought to function as an epigenetic mark that specifies centromere identity. Here we identify the essential centromere protein CENP-N as the first protein to selectively bind CENP-A nucleosomes but not H3 nucleosomes. CENP-N bound CENP-A nucleosomes in a DNA sequence-independent manner, but did not bind soluble CENP-A-H4 tetramers. Mutations in CENP-N that reduced its affinity for CENP-A nucleosomes caused defects in CENP-N localization and had dominant effects on the recruitment of CENP-H, CENP-I and CENP-K to centromeres. Depletion of CENP-N using siRNA (short interfering RNA) led to similar centromere assembly defects and resulted in reduced assembly of nascent CENP-A into centromeric chromatin. These data suggest that CENP-N interprets the information encoded within CENP-A nucleosomes and recruits other proteins to centromeric chromatin that are required for centromere function and propagation.
Figures
Comment in
-
A reader for centromeric chromatin.Nat Cell Biol. 2009 Jul;11(7):793-5. doi: 10.1038/ncb0709-793. Nat Cell Biol. 2009. PMID: 19568265
References
-
- Cheeseman IM, Desai A. Molecular architecture of the kinetochore-microtubule interface. Nat Rev Mol Cell Biol. 2008;9:33–46. - PubMed
-
- Choo KH. Domain Organization at the Centromere and Neocentromere. Dev Cell. 2001;1:165–177. - PubMed
-
- Carroll CW, Straight AF. Centromere formation: from epigenetics to self-assembly. Trends Cell Biol. 2006;16:70–78. - PubMed
-
- Okada M, et al. The CENP-H-I complex is required for the efficient incorporation of newly synthesized CENP-A into centromeres. Nat Cell Biol. 2006;8:446–457. - PubMed
-
- Foltz DR, et al. The human CENP-A centromeric nucleosome-associated complex. Nat Cell Biol. 2006;8:458–469. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
