

The HaDREB2 transcription factor enhances basal thermotolerance and longevity of seeds through functional interaction with HaHSFA9
- PMID: 19545370
- PMCID: PMC2706249
- DOI: 10.1186/1471-2229-9-75
The HaDREB2 transcription factor enhances basal thermotolerance and longevity of seeds through functional interaction with HaHSFA9
Abstract
Background: Transcription factor HaDREB2 was identified in sunflower (Helianthus annuus L.) as a drought-responsive element-binding factor 2 (DREB2) with unique properties. HaDREB2 and the sunflower Heat Shock Factor A9 (HaHSFA9) co-activated the Hahsp17.6G1 promoter in sunflower embryos. Both factors could be involved in transcriptional co-activation of additional small heat stress protein (sHSP) promoters, and thus contribute to the HaHSFA9-mediated enhancement of longevity and basal thermotolerance of seeds.
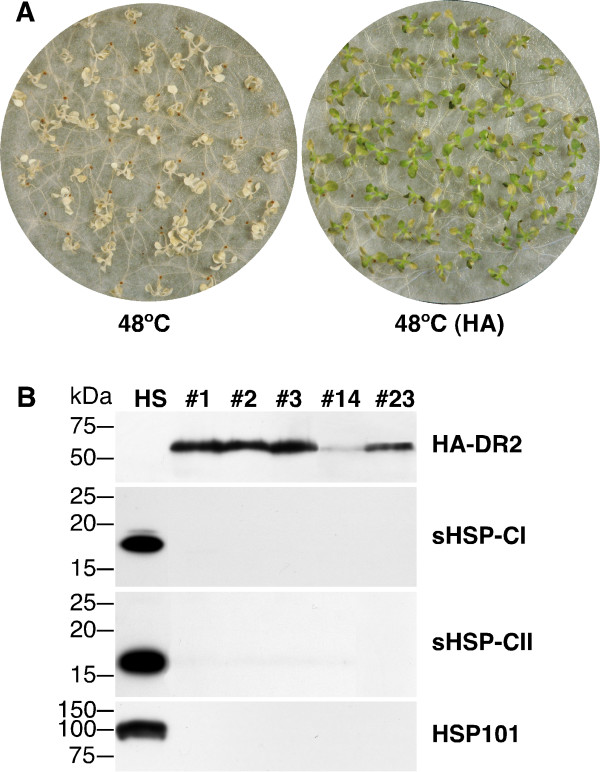
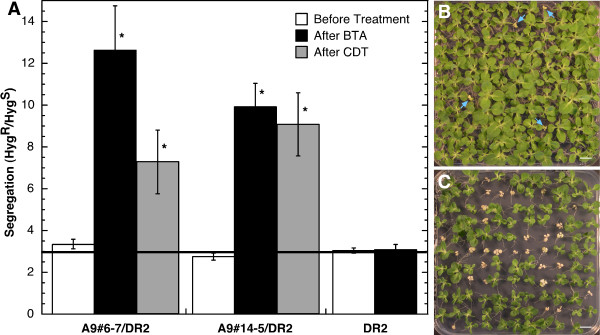
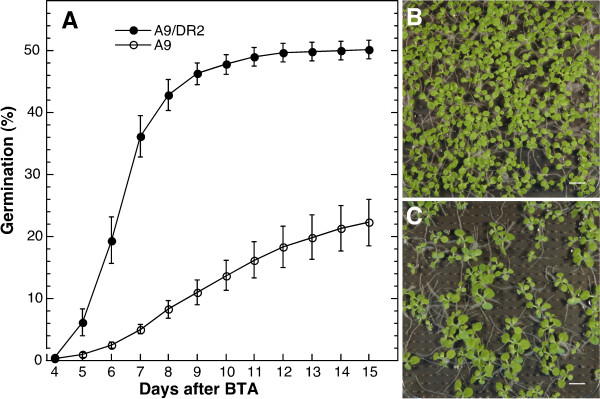
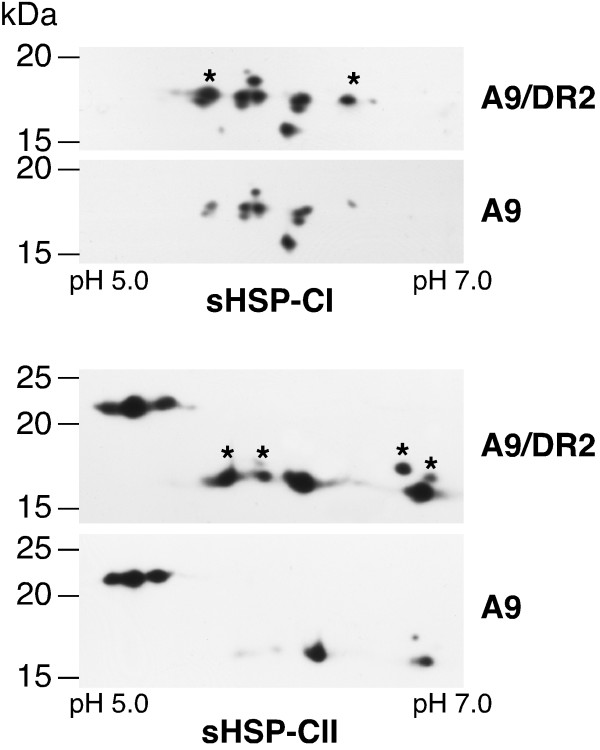
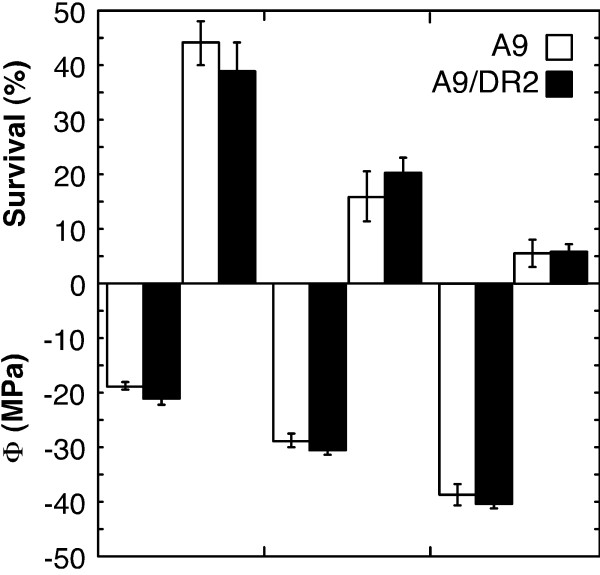
Results: We found that overexpression of HaDREB2 in seeds did not enhance longevity. This was deduced from assays of basal thermotolerance and controlled seed-deterioration, which were performed with transgenic tobacco. Furthermore, the constitutive overexpression of HaDREB2 did not increase thermotolerance in seedlings or result in the accumulation of HSPs at normal growth temperatures. In contrast, when HaDREB2 and HaHSFA9 were conjointly overexpressed in seeds, we observed positive effects on seed longevity, beyond those observed with overexpression of HaHSFA9 alone. Such additional effects are accompanied by a subtle enhancement of the accumulation of subsets of sHSPs belonging to the CI and CII cytosolic classes.
Conclusion: Our results reveal the functional interdependency of HaDREB2 and HaHSFA9 in seeds. HaDREB2 differs from other previously characterized DREB2 factors in plants in terms of its unique functional interaction with the seed-specific HaHSFA9 factor. No functional interaction between HaDREB2 and HaHSFA9 was observed when both factors were conjointly overexpressed in vegetative tissues. We therefore suggest that additional, seed-specific factors, or protein modifications, could be required for the functional interaction between HaDREB2 and HaHSFA9.
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
