

Desmoglein 1-dependent suppression of EGFR signaling promotes epidermal differentiation and morphogenesis
- PMID: 19546243
- PMCID: PMC2712955
- DOI: 10.1083/jcb.200809044
Desmoglein 1-dependent suppression of EGFR signaling promotes epidermal differentiation and morphogenesis
Abstract
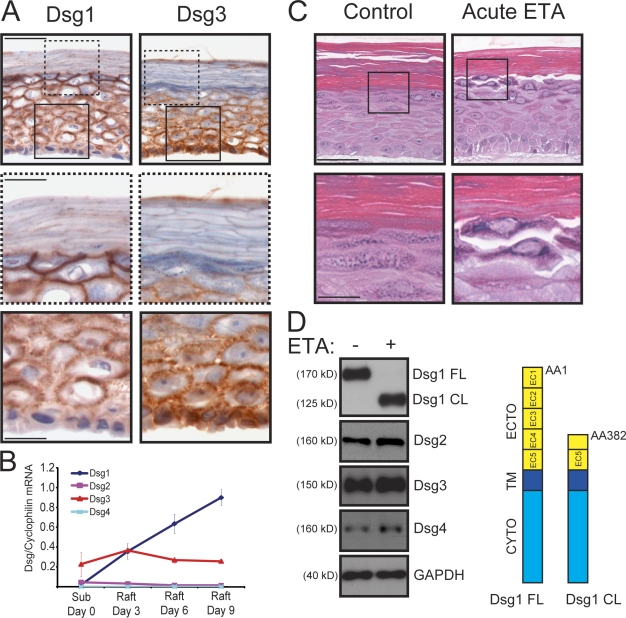
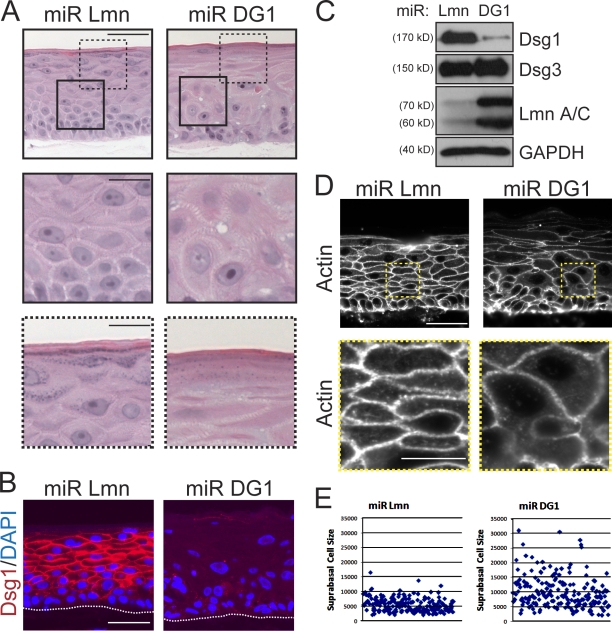
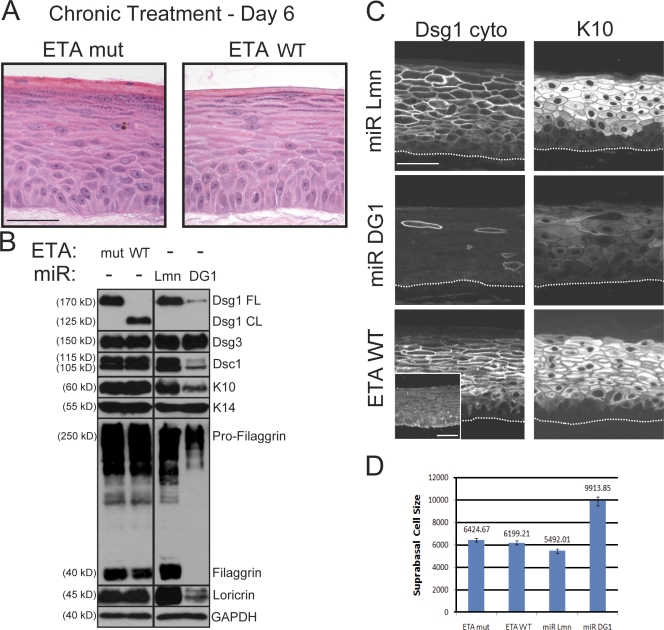
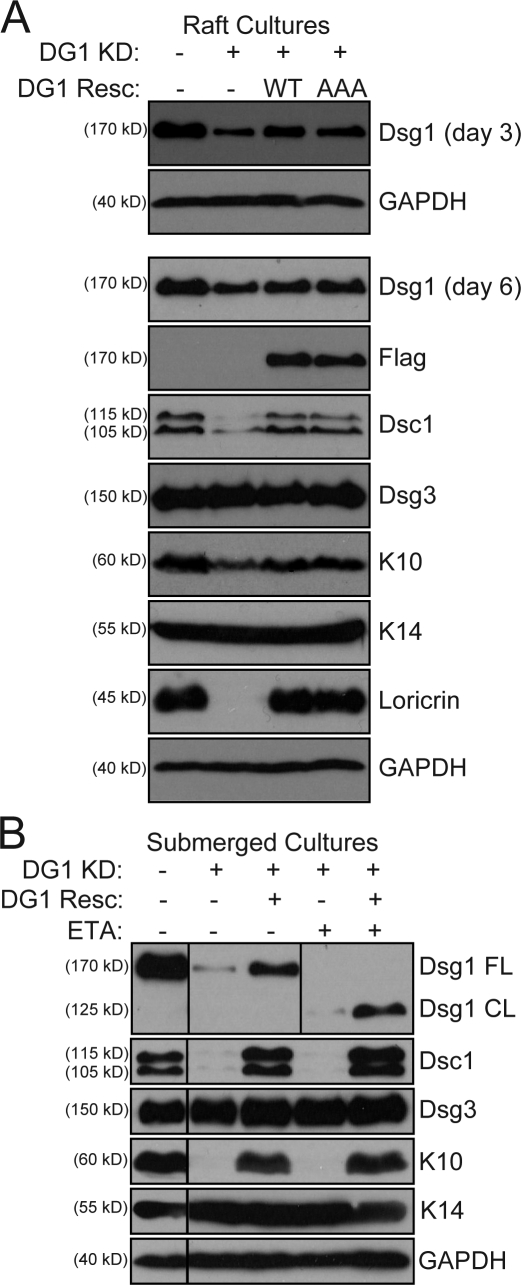
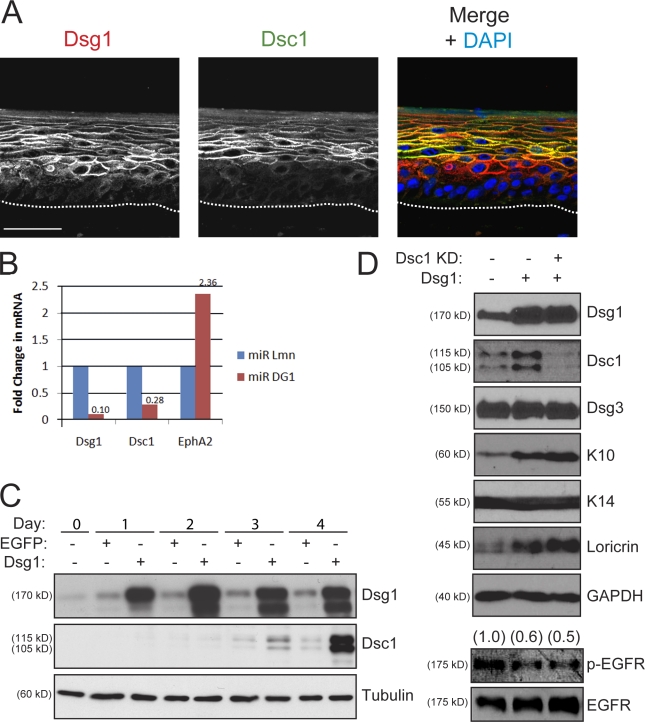
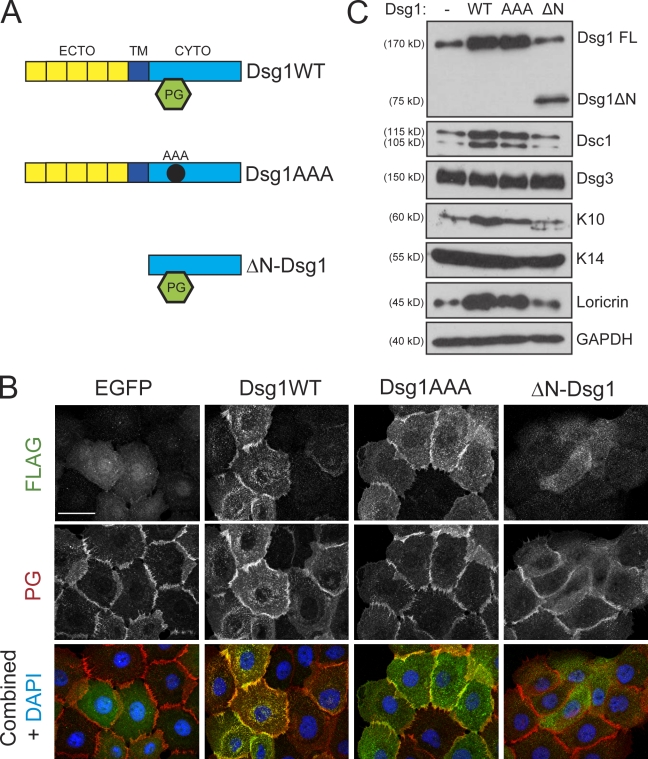
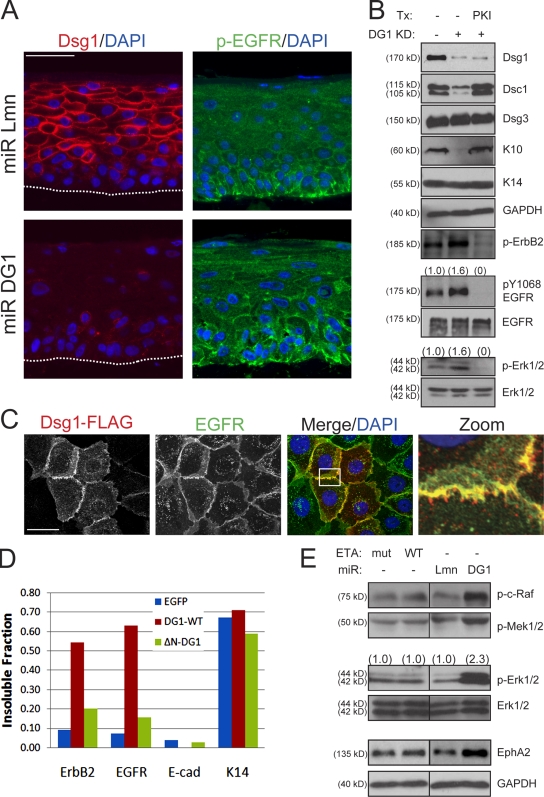
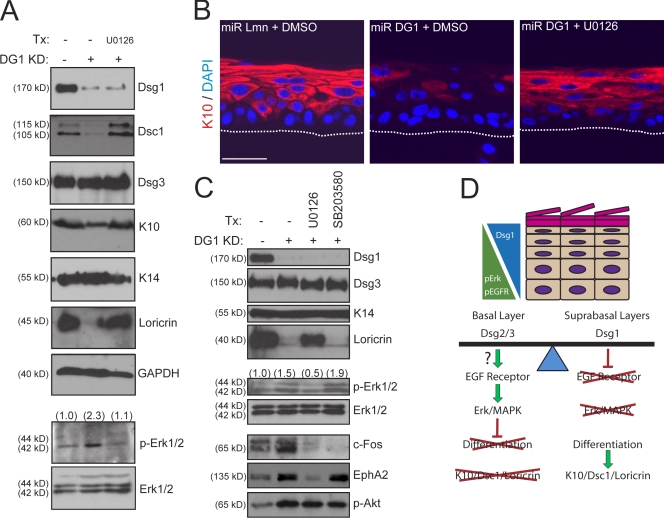
Dsg1 (desmoglein 1) is a member of the cadherin family of Ca(2+)-dependent cell adhesion molecules that is first expressed in the epidermis as keratinocytes transit out of the basal layer and becomes concentrated in the uppermost cell layers of this stratified epithelium. In this study, we show that Dsg1 is not only required for maintaining epidermal tissue integrity in the superficial layers but also supports keratinocyte differentiation and suprabasal morphogenesis. Dsg1 lacking N-terminal ectodomain residues required for adhesion remained capable of promoting keratinocyte differentiation. Moreover, this capability did not depend on cytodomain interactions with the armadillo protein plakoglobin or coexpression of its companion suprabasal cadherin, Dsc1 (desmocollin 1). Instead, Dsg1 was required for suppression of epidermal growth factor receptor-Erk1/2 (extracellular signal-regulated kinase 1/2) signaling, thereby facilitating keratinocyte progression through a terminal differentiation program. In addition to serving as a rigid anchor between adjacent cells, this study implicates desmosomal cadherins as key components of a signaling axis governing epithelial morphogenesis.
Figures
References
-
- Amagai M., Matsuyoshi N., Wang Z.H., Andl C., Stanley J.R. 2000. Toxin in bullous impetigo and staphylococcal scalded-skin syndrome targets desmoglein 1.Nat. Med. 6:1275–1277 - PubMed
-
- Arnemann J., Sullivan K.H., Magee A.I., King I.A., Buxton R.S. 1993. Stratification-related expression of isoforms of the desmosomal cadherins in human epidermis.J. Cell Sci. 104:741–750 - PubMed
-
- Bailleul B., Surani M.A., White S., Barton S.C., Brown K., Blessing M., Jorcano J., Balmain A. 1990. Skin hyperkeratosis and papilloma formation in transgenic mice expressing a ras oncogene from a suprabasal keratin promoter.Cell. 62:697–708 - PubMed
-
- Bazzi H., Getz A., Mahoney M.G., Ishida-Yamamoto A., Langbein L., Wahl J.K., III, Christiano A.M. 2006. Desmoglein 4 is expressed in highly differentiated keratinocytes and trichocytes in human epidermis and hair follicle.Differentiation. 74:129–140 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
