

Review
doi: 10.1038/nrm2718.
Boveri revisited: chromosomal instability, aneuploidy and tumorigenesis
Affiliations
- PMID: 19546858
- PMCID: PMC3154738
- DOI: 10.1038/nrm2718
Item in Clipboard
Review
Boveri revisited: chromosomal instability, aneuploidy and tumorigenesis
Nat Rev Mol Cell Biol.
2009 Jul.
Abstract
The mitotic checkpoint is a major cell cycle control mechanism that guards against chromosome missegregation and the subsequent production of aneuploid daughter cells. Most cancer cells are aneuploid and frequently missegregate chromosomes during mitosis. Indeed, aneuploidy is a common characteristic of tumours, and, for over 100 years, it has been proposed to drive tumour progression. However, recent evidence has revealed that although aneuploidy can increase the potential for cellular transformation, it also acts to antagonize tumorigenesis in certain genetic contexts. A clearer understanding of the tumour suppressive function of aneuploidy might reveal new avenues for anticancer therapy.
Figures
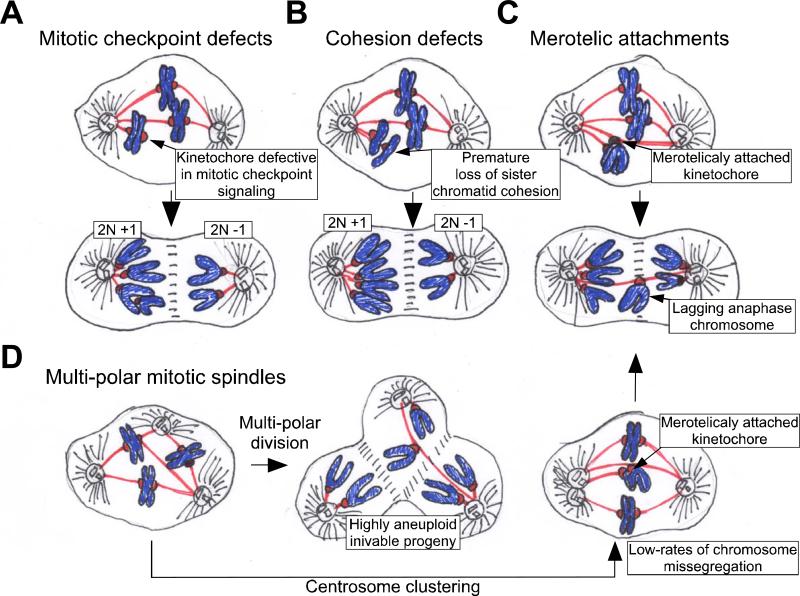
There are several pathways by which cells may gain or lose chromosomes during mitosis. A. Defects in mitotic checkpoint signaling. A weakened mitotic checkpoint may allow cells to enter anaphase in the presence of unattached or misaligned chromosomes. As a consequence, both copies of one chromosome may be deposited into a single daughter cell. B. Cohesion defects. If sister chromatid cohesion is lost prematurely or persists during anaphase then chromosomes may be missegregated. C. Merotelic attachment. One kinetochore may attach to microtubules from both poles of the spindle. If these attachments persist into anaphase then lagging chromatid pairs may be missegregated or excluded from both daughter cells during cytokinesis. D. Multipolar mitotic divisions. Cells possessing more than two centrosomes may form multiple spindle poles during mitosis. If this defect is not corrected, then a multipolar division will occur resulting in the production of highly aneuploid and often inviable daughter cells. Often, however, centrosomes in multipolar spindles cluster into two groups to allow cells to divide in a bipolar fashion. Centrosome clustering will increase the formation of incorrect kinetocore-microtubule attachments (such as merotelic attachments). Extra centrosomes are thus, capable of driving low rates of chromosome missegregation through a mechanism independent of multipolar divisions.
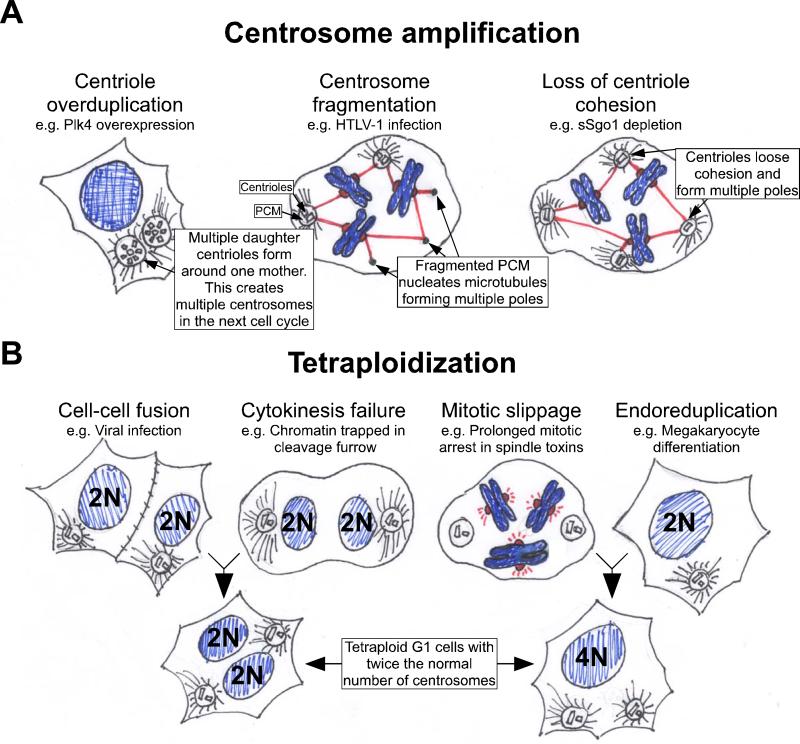
The centrosome consists of a pair of connected centrioles surrounded by the pericentriolar material (PCM). There are two major mechanisms by which cells can gain extra centrosomes. A. Centrosome amplification. Defects in the processes controlling centriole replication can lead to centriole overduplication, which results in multiple centrosomes in the next cell cycle. This process can occur when PLK4, a regulator of centriole biogenesis, is overexpressed, . Impairment of centrosome structure can cause fragmentation of the pericentriolar material. The acentriolar fragments can then serve to nucleate microtubules and create multipolar spindles. This has been found to occur following cellular infection with the human T cell lymphotrophic virus Type 1 (HTLV-1). Finally, defects in centriole cohesion can lead to the separation of paired centrioles before the completion of chromosome segregation, creating multiple microtubule nucleating foci. Cells with reduced levels of sSgo1 have been shown to loose centriole cohesion prematurely. B. Cells become tetraploid. This can occur following cell-cell fusion or after cytokinesis failure. Alternatively, cells may skip mitosis altogether and endoreduplicate or “slip” out of mitosis and progress into the next cell cycle without undergoing anaphase or cytokinesis. In all these situations G1 tetraploid cells are created with two centrosomes that are duplicated during the next cell cycle.
References
-
- Boveri T. Ueber mehrpolige Mitosen als Mittel zur Analyse des Zellkerns. Verh Phys-med Ges Würzburg NF. 1902;35:67–90.
-
- Hansemann D. Ueber asymmetrische Zelltheilung in Epithelkrebsen und deren biologische Bedeutung. Arch Pathol Anat Physiol Klin Medicin. 1980;119:299–326.
-
- Boveri T. Zur Frage der Entstehung maligner Tumoren. Fischer, Jena. 1914
-
- Weaver BA, Cleveland DW. Does aneuploidy cause cancer? Curr Opin Cell Biol. 2006;18:658–67. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
