

The current structural and functional understanding of APOBEC deaminases
- PMID: 19547914
- PMCID: PMC11115857
- DOI: 10.1007/s00018-009-0070-y
The current structural and functional understanding of APOBEC deaminases
Abstract
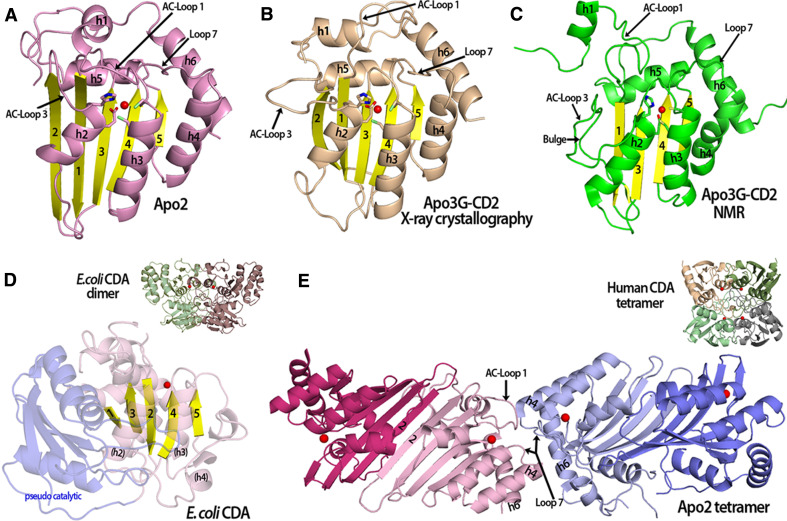
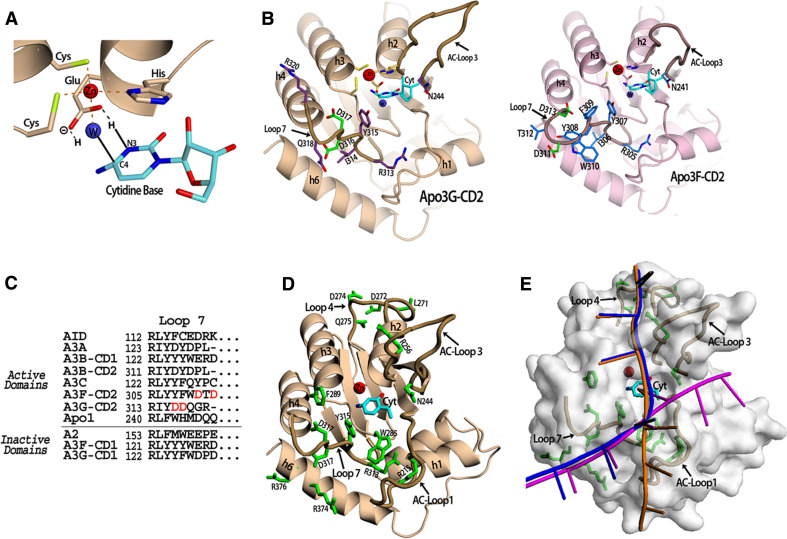
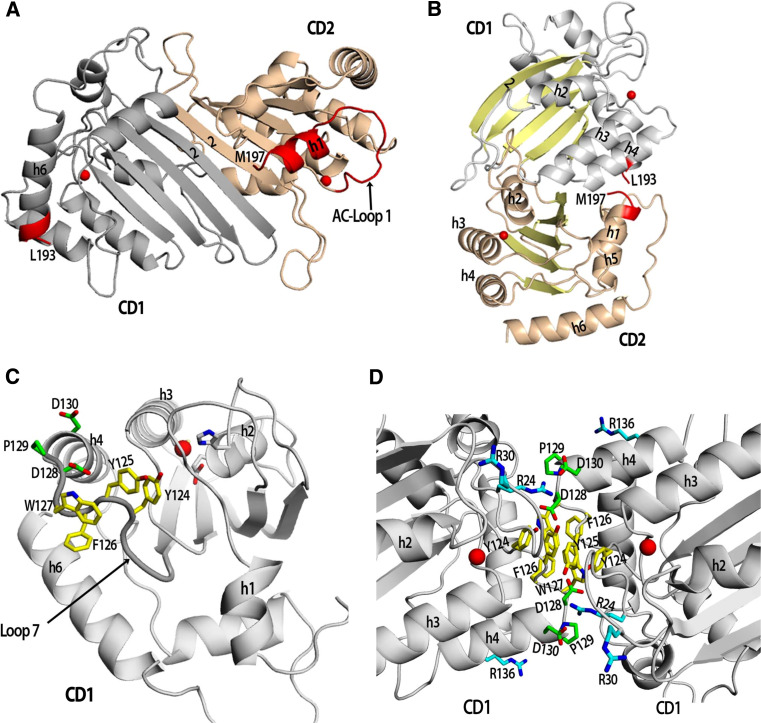
The apolipoprotein B mRNA-editing enzyme catalytic polypeptide (APOBEC) family of cytidine deaminases has emerged as an intensively studied field as a result of their important biological functions. These enzymes are involved in lipid metabolism, antibody diversification, and the inhibition of retrotransposons, retroviruses, and some DNA viruses. The APOBEC proteins function in these roles by deaminating single-stranded (ss) DNA or RNA. There are two high-resolution crystal structures available for the APOBEC family, Apo2 and the C-terminal catalytic domain (CD2) of Apo3G or Apo3G-CD2 [Holden et al. (Nature 456:121-124, 2008); Prochnow et al. (Nature 445:447-451, 2007)]. Additionally, the structure of Apo3G-CD2 has also been determined using NMR [Chen et al. (Nature 452:116-119, 2008); Furukawa et al. (EMBO J 28:440-451, 2009); Harjes et al. (J Mol Biol, 2009)]. A detailed structural analysis of the APOBEC proteins and a comparison to other zinc-coordinating deaminases can facilitate our understanding of how APOBEC proteins bind nucleic acids, recognize substrates, and form oligomers. Here, we review the recent development of structural and functional studies that apply to Apo3G as well as the APOBEC deaminase family.
Figures
References
-
- Prochnow C, Bransteitter R, Klein MG, Goodman MF, Chen XS. The APOBEC-2 crystal structure and functional implications for the deaminase AID. Nature. 2007;445:447–451. - PubMed
-
- Chen KM, Harjes E, Gross PJ, Fahmy A, Lu Y, Shindo K, Harris RS, Matsuo H. Structure of the DNA deaminase domain of the HIV-1 restriction factor APOBEC3G. Nature. 2008;452:116–119. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
