

Transmitochondrial embryonic stem cells containing pathogenic mtDNA mutations are compromised in neuronal differentiation
- PMID: 19552636
- PMCID: PMC2730481
- DOI: 10.1111/j.1365-2184.2009.00612.x
Transmitochondrial embryonic stem cells containing pathogenic mtDNA mutations are compromised in neuronal differentiation
Abstract
Objectives: Defects of the mitochondrial genome (mtDNA) cause a series of rare, mainly neurological disorders. In addition, they have been implicated in more common forms of movement disorders, dementia and the ageing process. In order to try to model neuronal dysfunction associated with mitochondrial disease, we have attempted to establish a series of transmitochondrial mouse embryonic stem cells harbouring pathogenic mtDNA mutations.
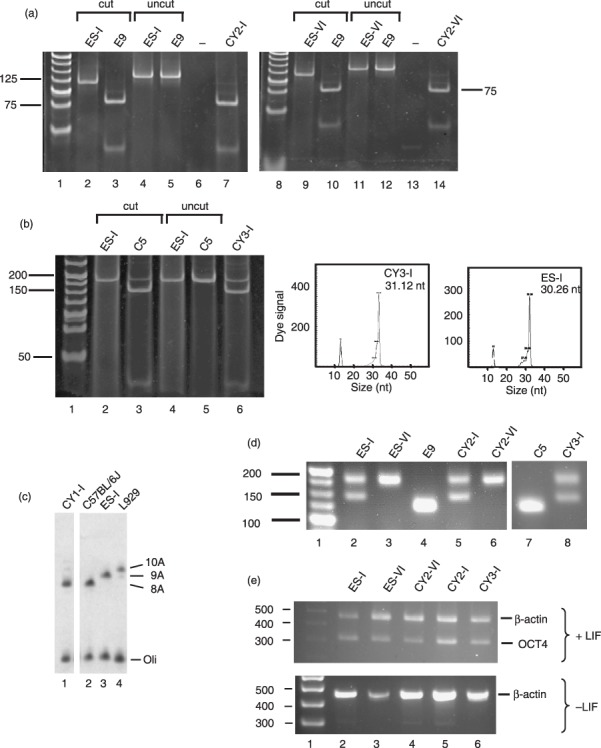
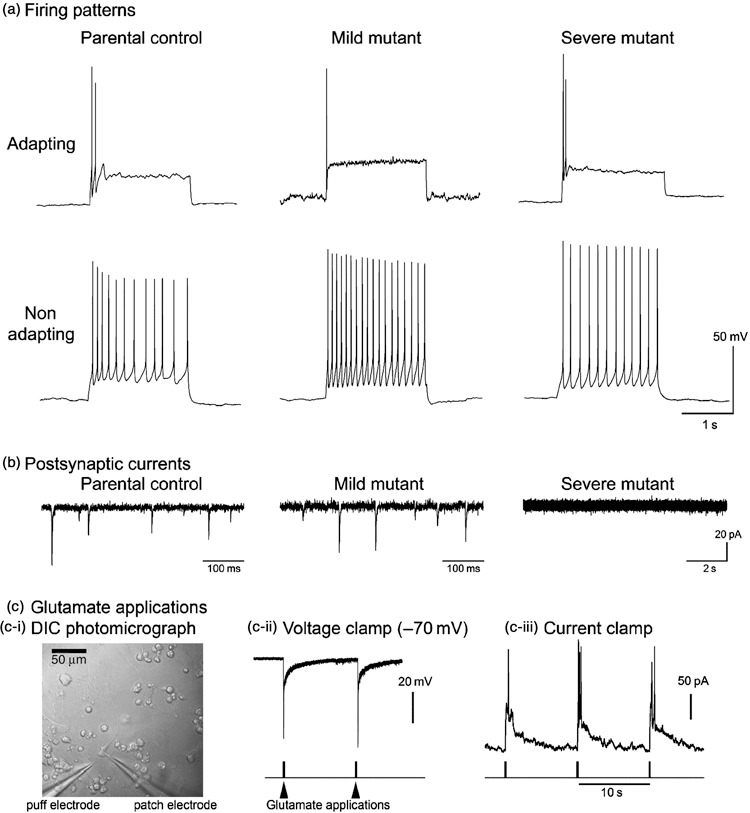
Materials and methods: Transmitochondrial embryonic stem cell cybrids were generated by fusion of cytoplasts carrying a variety of mtDNA mutations, into embryonic stem cells that had been pretreated with rhodamine 6G, to prevent transmission of endogenous mtDNA. Cybrids were differentiated into neurons and assessed for efficiency of differentiation and electrophysiological function.
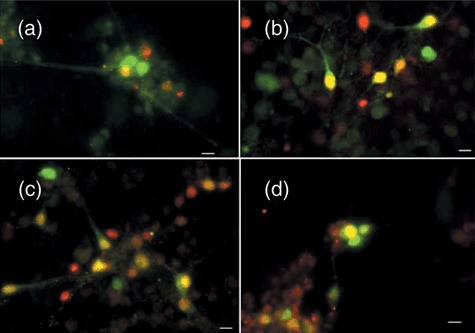
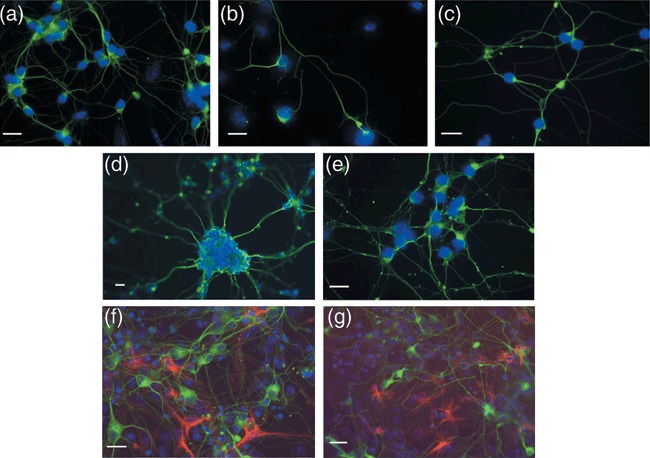
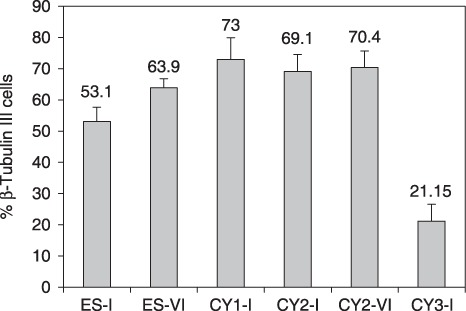
Results: Neuronal differentiation could occur, as indicated by expression of neuronal markers. Differentiation was impaired in embryonic stem cells carrying mtDNA mutations that caused severe biochemical deficiency. Electrophysiological tests showed evidence of synaptic activity in differentiated neurons carrying non-pathogenic mtDNA mutations or in those that caused a mild defect of respiratory activity. Again, however, neurons carrying mtDNA mutations that resulted in severe biochemical deficiency had marked reduction in post-synaptic events.
Conclusions: Differentiated neurons carrying severely pathogenic mtDNA defects can provide a useful model for understanding how such mutations can cause neuronal dysfunction.
Figures
References
-
- Bender A, Krishnan KJ, Morris CM, Taylor GA, Reeve AK, Perry RH et al (2006) High levels of mitochondrial DNA deletions in substantia nigra neurons in aging and Parkinson disease. Nat. Genet. 38, 515–517. - PubMed
-
- Trifunovic A, Wredenberg A, Falkenberg M, Spelbrink JN, Rovio AT, Bruder CE et al (2004) Premature ageing in mice expressing defective mitochondrial DNA polymerase. Nature 429, 417–423. - PubMed
-
- Vermulst M, Wanagat J, Kujoth GC, Bielas JH, Rabinovitch PS, Prolla TA et al (2008) DNA deletions and clonal mutations drive premature aging in mitochondrial mutator mice. Nat. Genet. 40, 392–394. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
