

JNK/AP-1 pathway is involved in tumor necrosis factor-alpha induced expression of vascular endothelial growth factor in MCF7 cells
- PMID: 19553068
- PMCID: PMC3449307
- DOI: 10.1016/j.biopha.2009.04.045
JNK/AP-1 pathway is involved in tumor necrosis factor-alpha induced expression of vascular endothelial growth factor in MCF7 cells
Abstract
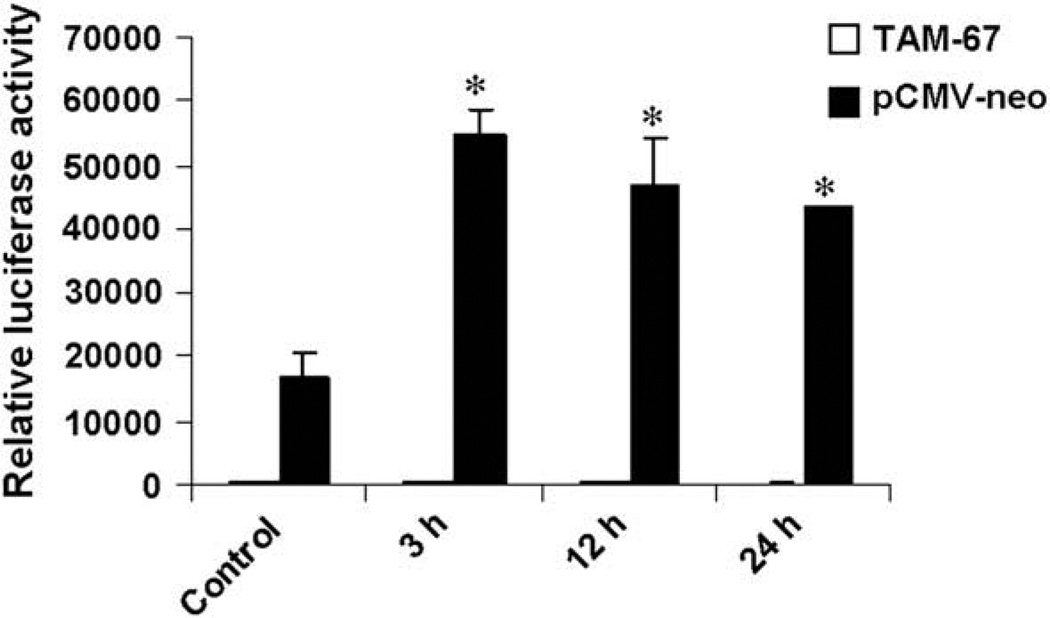
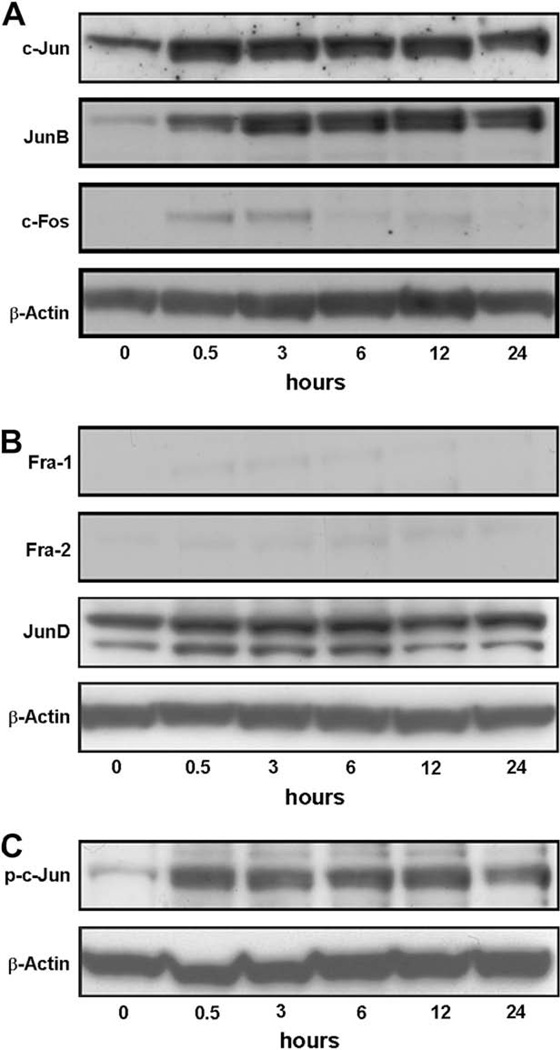
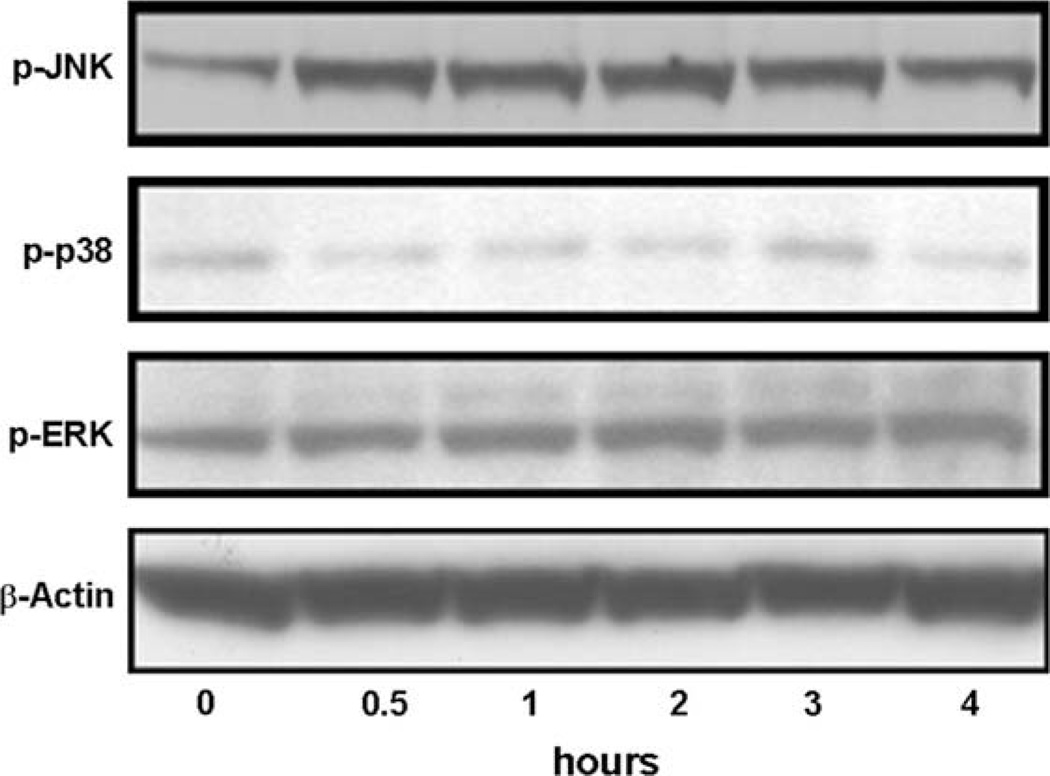
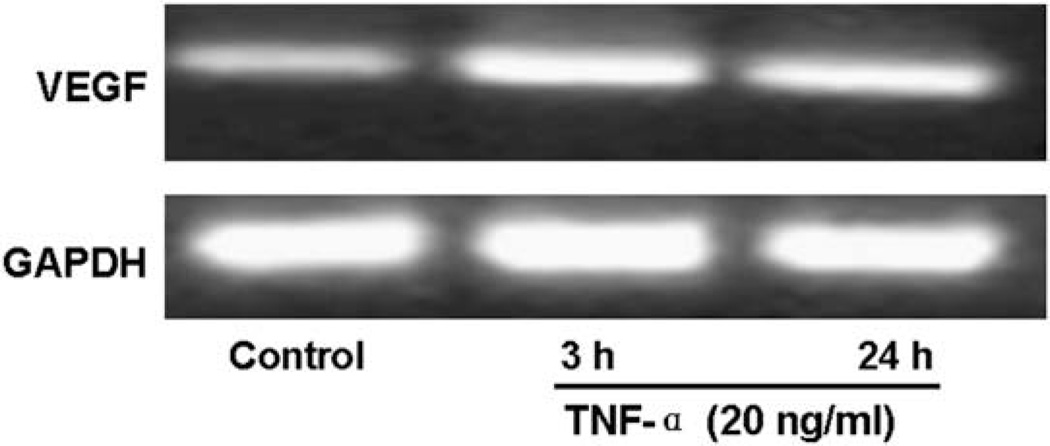
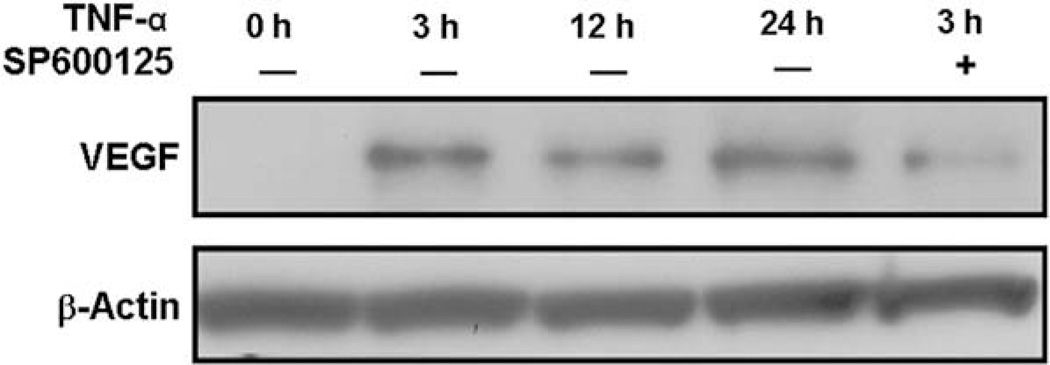
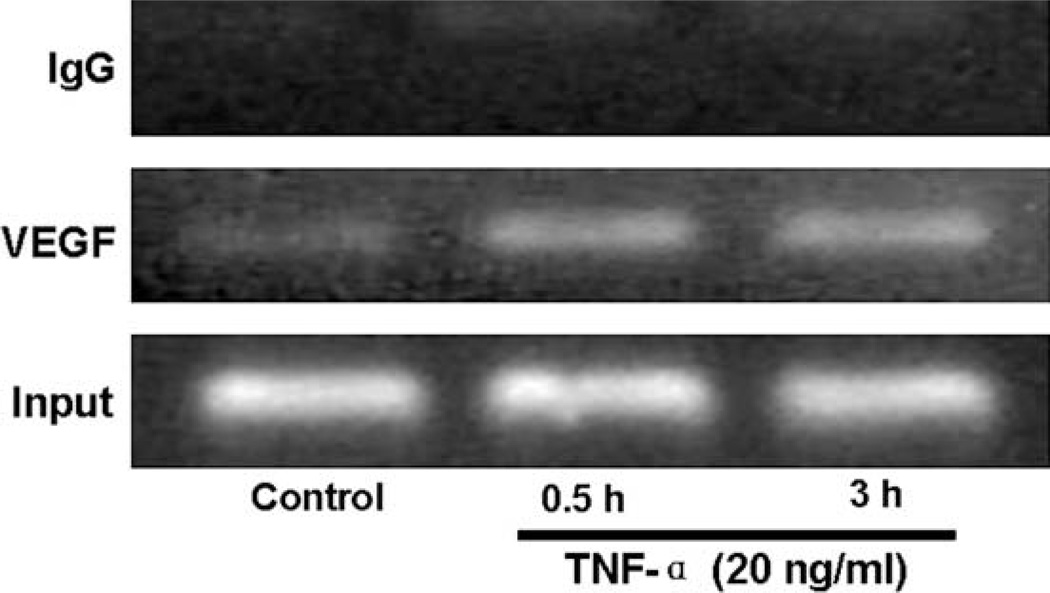
Vascular endothelial growth factor (VEGF) has been implicated in breast tumor angiogenesis. And tumor necrosis factor-alpha (TNF-alpha) is a positive regulator of VEGF. This study was aimed to identify the signalling pathway of TNF-alpha in VEGF expression regulation in breast cancer cell line MCF7. Using luciferase reporter assays, we demonstrated that TNF-alpha significantly increased activator protein-1 (AP-1) transcriptional activity in the MCF7 cells. The expression of the AP-1 family members c-Jun, c-Fos and JunB and phosphorylation levels of c-Jun were upregulated by TNF-alpha, whereas other AP-1 family members Fra-1, Fra-2, and JunD were unaffected. The activation of AP-1 was associated with the formation of p-c-Jun-c-Jun and p-c-Jun-JunB homodimers. Furthermore, the phosphorylation levels of c-Jun N-terminal kinase (JNK) but not P38 and ERK were elevated by TNF-alpha in MCF7 cells. TNF-alpha potently upregulated the mRNA and protein levels of VEGF, which were significantly reversed by JNK inhibitor SP600125. Finally using chromatin immunoprecipitation (CHIP) assays, we found that p-c-Jun bound to the VEGF promoter and regulated VEGF transcription directly. These data suggest that the pro-inflammatory cytokine TNF-alpha is a critical regulator of VEGF expression in breast cancer cells, at least partially via a JNK and AP-1 dependent pathway.
Figures


References
-
- Ferrara N, Davis-Smyth T. The biology of vascular endothelial growth factor. Endocr Rev. 1997;18(1):4–25. - PubMed
-
- Leek RD, Harris AL, Lewis CE. Cytokine networks in solid human tumors: regulation of angiogenesis. J Leukoc Biol. 1994;56(4):423–435. - PubMed
-
- Tischer E, Mitchell R, Hartman T, Silva M, Gospodarowicz D, Fiddes JC, et al. The human gene for vascular endothelial growth factor. Multiple protein forms are encoded through alternative exon splicing. J Biol Chem. 1991;266(18):11947–11954. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Research Materials
Miscellaneous
