

Antibody specificities associated with neutralization breadth in plasma from human immunodeficiency virus type 1 subtype C-infected blood donors
- PMID: 19553335
- PMCID: PMC2738176
- DOI: 10.1128/JVI.00758-09
Antibody specificities associated with neutralization breadth in plasma from human immunodeficiency virus type 1 subtype C-infected blood donors
Abstract
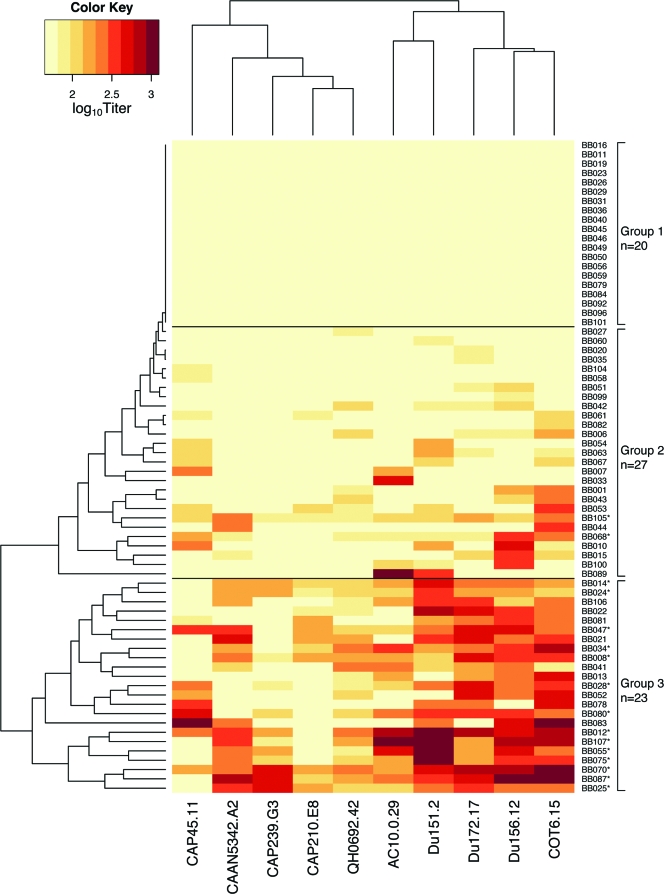
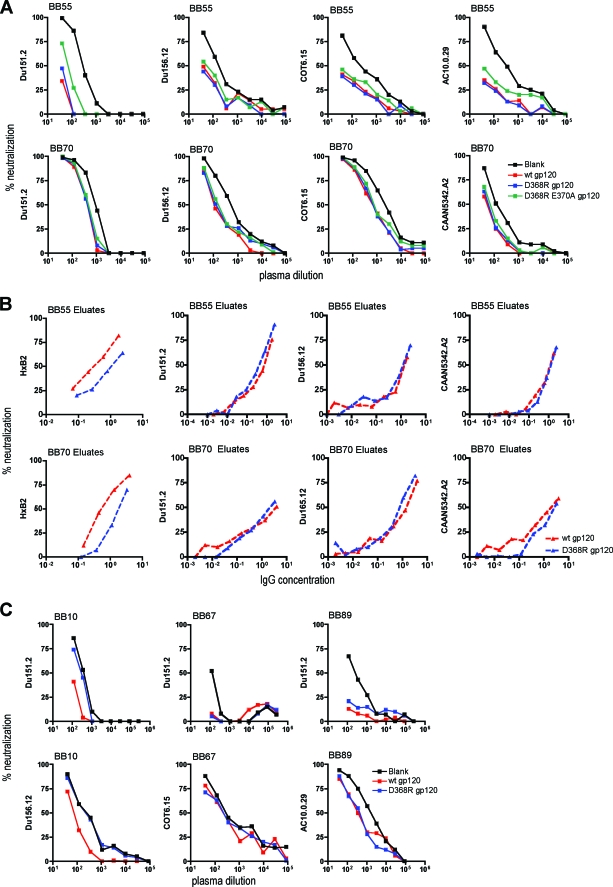
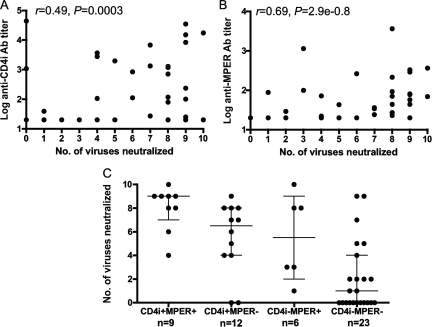
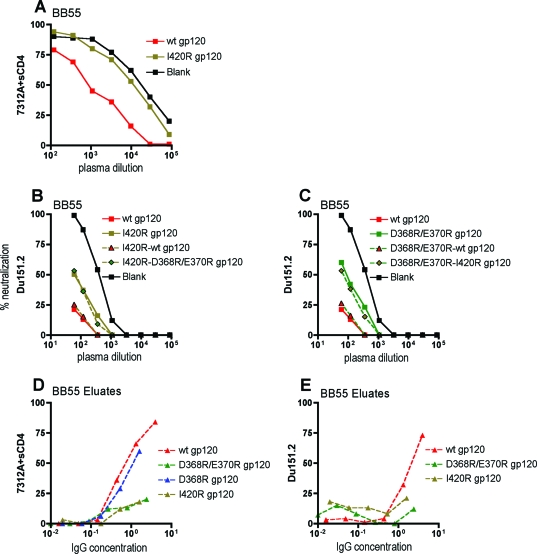
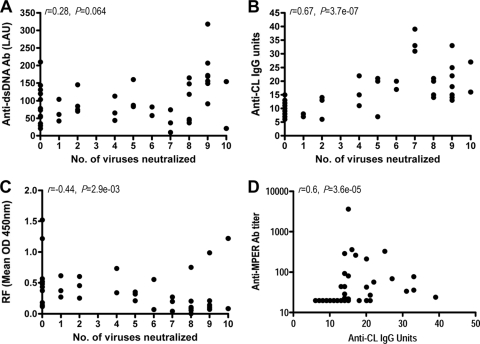
Defining the specificities of the anti-human immunodeficiency virus type 1 (HIV-1) envelope antibodies able to mediate broad heterologous neutralization will assist in identifying targets for an HIV-1 vaccine. We screened 70 plasmas from chronically HIV-1-infected individuals for neutralization breadth. Of these, 16 (23%) were found to neutralize 80% or more of the viruses tested. Anti-CD4 binding site (CD4bs) antibodies were found in almost all plasmas independent of their neutralization breadth, but they mainly mediated neutralization of the laboratory strain HxB2 with little effect on the primary virus, Du151. Adsorption with Du151 monomeric gp120 reduced neutralizing activity to some extent in most plasma samples when tested against the matched virus, although these antibodies did not always confer cross-neutralization. For one plasma, this activity was mapped to a site overlapping the CD4-induced (CD4i) epitope and CD4bs. Anti-membrane-proximal external region (MPER) (r = 0.69; P < 0.001) and anti-CD4i (r = 0.49; P < 0.001) antibody titers were found to be correlated with the neutralization breadth. These anti-MPER antibodies were not 4E10- or 2F5-like but spanned the 4E10 epitope. Furthermore, we found that anti-cardiolipin antibodies were correlated with the neutralization breadth (r = 0.67; P < 0.001) and anti-MPER antibodies (r = 0.6; P < 0.001). Our study suggests that more than one epitope on the envelope glycoprotein is involved in the cross-reactive neutralization elicited during natural HIV-1 infection, many of which are yet to be determined, and that polyreactive antibodies are possibly involved in this phenomenon.
Figures
References
-
- Binley, J. M., E. A. Lybarger, E. T. Crooks, M. S. Seaman, E. Gray, K. L. Davis, J. M. Decker, D. Wycuff, L. Harris, N. Hawkins, B. Wood, C. Nathe, D. Richman, G. D. Tomaras, F. Bibollet-Ruche, J. E. Robinson, L. Morris, G. M. Shaw, D. C. Montefiori, and J. R. Mascola. 2008. Profiling the specificity of neutralizing antibodies in a large panel of plasmas from patients chronically infected with human immunodeficiency virus type 1 subtypes B and C. J. Virol. 8211651-11668. - PMC - PubMed
-
- Binley, J. M., R. W. Sanders, B. Clas, N. Schuelke, A. Master, Y. Guo, F. Kajumo, D. J. Anselma, P. J. Maddon, W. C. Olson, and J. P. Moore. 2000. A recombinant human immunodeficiency virus type 1 envelope glycoprotein complex stabilized by an intermolecular disulfide bond between the gp120 and gp41 subunits is an antigenic mimic of the trimeric virion-associated structure. J. Virol. 74627-643. - PMC - PubMed
-
- Binley, J. M., T. Wrin, B. Korber, M. B. Zwick, M. Wang, C. Chappey, G. Stiegler, R. Kunert, S. Zolla-Pazner, H. Katinger, C. J. Petropoulos, and D. R. Burton. 2004. Comprehensive cross-clade neutralization analysis of a panel of anti-human immunodeficiency virus type 1 monoclonal antibodies. J. Virol. 7813232-13252. - PMC - PubMed
-
- Bonnet, F., J. J. Pineau, J. L. Taupin, A. Feyler, M. Bonarek, S. de Witte, N. Bernard, D. Lacoste, P. Morlat, and J. Beylot. 2003. Prevalence of cryoglobulinemia and serological markers of autoimmunity in human immunodeficiency virus infected individuals: a cross-sectional study of 97 patients. J. Rheumatol. 302005-2010. - PubMed
-
- Braibant, M., S. Brunet, D. Costagliola, C. Rouzioux, H. Agut, H. Katinger, B. Autran, and F. Barin. 2006. Antibodies to conserved epitopes of the HIV-1 envelope in sera from long-term non-progressors: prevalence and association with neutralizing activity. AIDS 201923-1930. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
