

Remobilization of leaf S compounds and senescence in response to restricted sulphate supply during the vegetative stage of oilseed rape are affected by mineral N availability
- PMID: 19553370
- PMCID: PMC2718225
- DOI: 10.1093/jxb/erp172
Remobilization of leaf S compounds and senescence in response to restricted sulphate supply during the vegetative stage of oilseed rape are affected by mineral N availability
Abstract
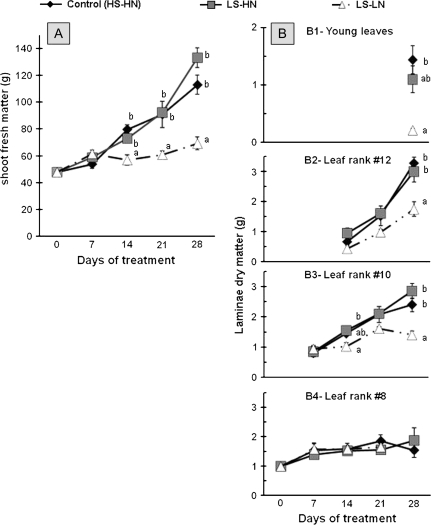
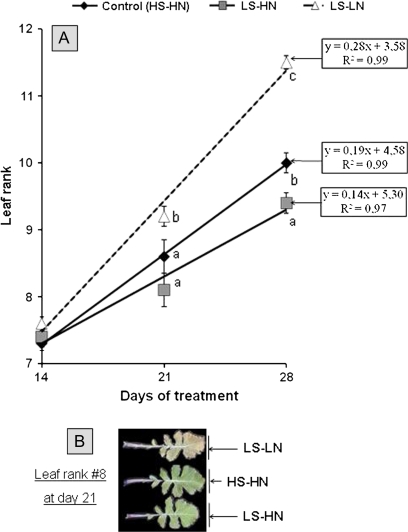
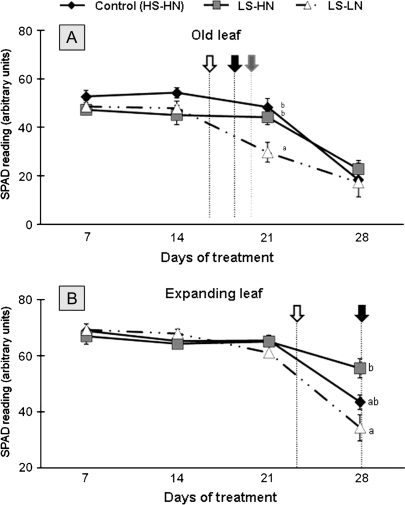
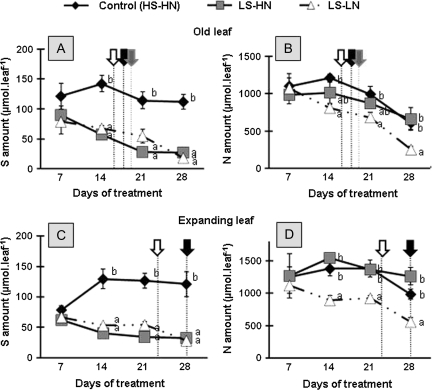
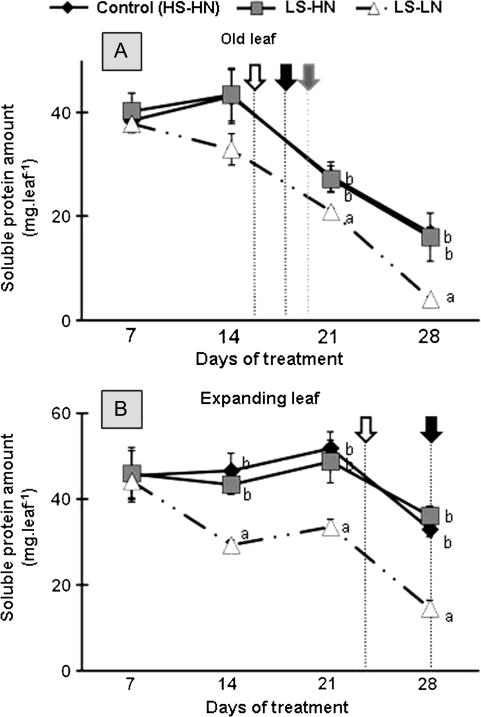
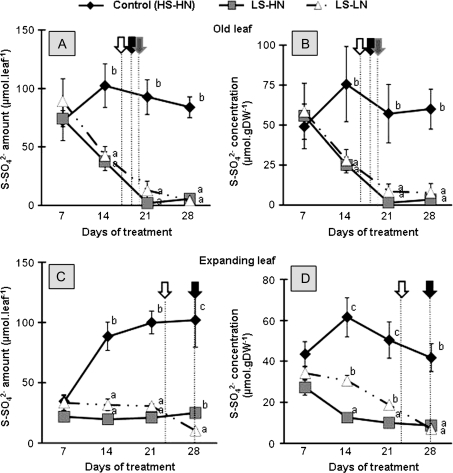
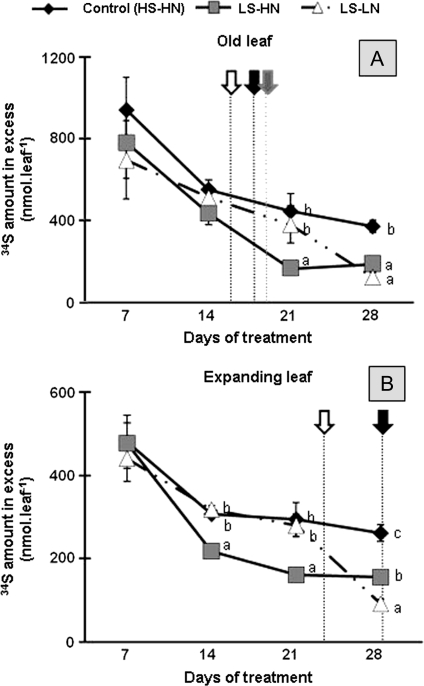
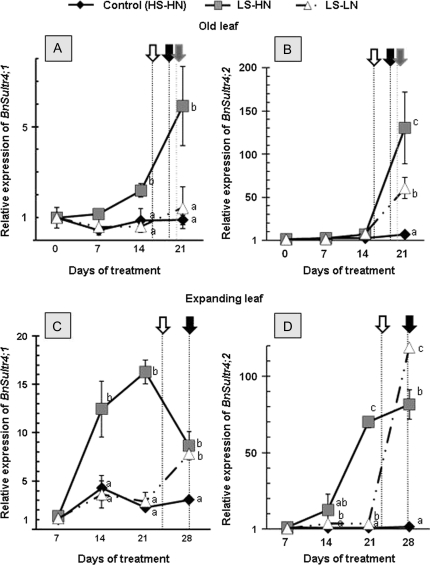
The impact of sulphur limitation on the remobilization of endogenous S compounds during the rosette stage of oilseed rape, and the interactions with N availability on these processes, were examined using a long-term (34)SO(4)(2-) labelling method combined with a study of leaf senescence progression (using SAG12/Cab as a molecular indicator) and gene expression of the transporters, BnSultr4;1 and BnSultr4;2, involved in vacuolar sulphate efflux. After 51 d on hydroponic culture at 0.3 mM (34)SO(4)(2-) (1 atom% excess), the labelling was stopped and plants were subject for 28 d to High S-High N (HS-HN, control), Low S-High N (LS-HN) or Low S-Low N (LS-LN) conditions. Compared with the control, LS-HN plants showed delayed leaf senescence and, whilst the shoot growth and the foliar soluble protein amounts were not affected, S, (34)S, and SO(4)(2-) amounts in the old leaves declined rapidly and were associated with the up-regulation of BnSultr4;1. In LS-LN plants, shoot growth was reduced, leaf senescence was accelerated, and the rapid S mobilization in old leaves was accompanied by decreased (34)S and SO(4)(2-), higher protein mobilization, and up-regulation of BnSultr4;2, but without any change of expression of BnSultr4;1. The data suggest that to sustain the S demand for growth under S restriction (i) vacuolar SO(4)(2-) is specifically remobilized in LS-HN conditions without any acceleration of leaf senescence, (ii) SO(4)(2-) mobilization is related to an up-regulation of BnSultr4;1 and/or BnSultr4;2 expression, and (iii) the relationship between sulphate mobilization and up-regulation of expression of BnSultr4 genes is specifically dependent on the N availability.
Figures
References
-
- Barberon M, Berthomieu P, Clairotte M, Shibagaki N, Davidian JC, Gosti F. Unequal functional redundancy between the two Arabidopsis thaliana high-affinity sulphate transporters SULTR1;1 and SULTR1;2. New Phytologist. 2008;180:608–619. - PubMed
-
- Bradford MM. A rapid method for the quantitation of microgram quantities of protein utilizing the principle of protein–dye binding. Analytical Biochemistry. 1976;72:248–254. - PubMed
-
- Diaz C, Lemaître T, Christ A, Azzopardi M, Kato Y, Sato F, Morot-Gaudry JF, Le Dily F, Masclaux-Daubresse C. Nitrogen recycling and remobilization are differentially controlled by leaf senescence and development stage in Arabidopsis under low nitrogen nutrition. Plant Physiology. 2008;147:1437–1449. - PMC - PubMed
