

Evidence that a defective spindle assembly checkpoint is not the primary cause of maternal age-associated aneuploidy in mouse eggs
- PMID: 19553597
- PMCID: PMC2754889
- DOI: 10.1095/biolreprod.109.077909
Evidence that a defective spindle assembly checkpoint is not the primary cause of maternal age-associated aneuploidy in mouse eggs
Abstract
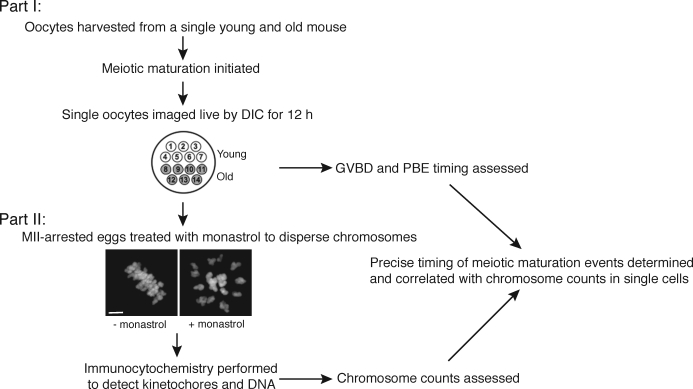
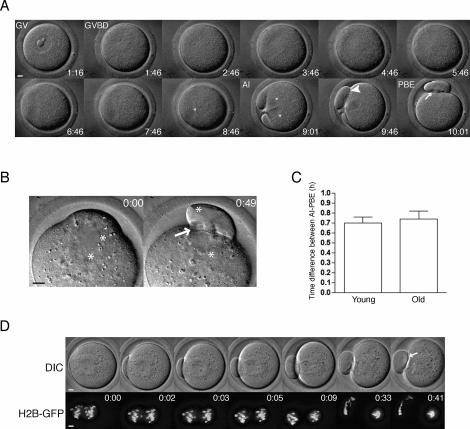
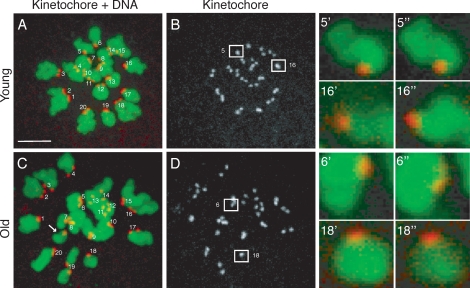
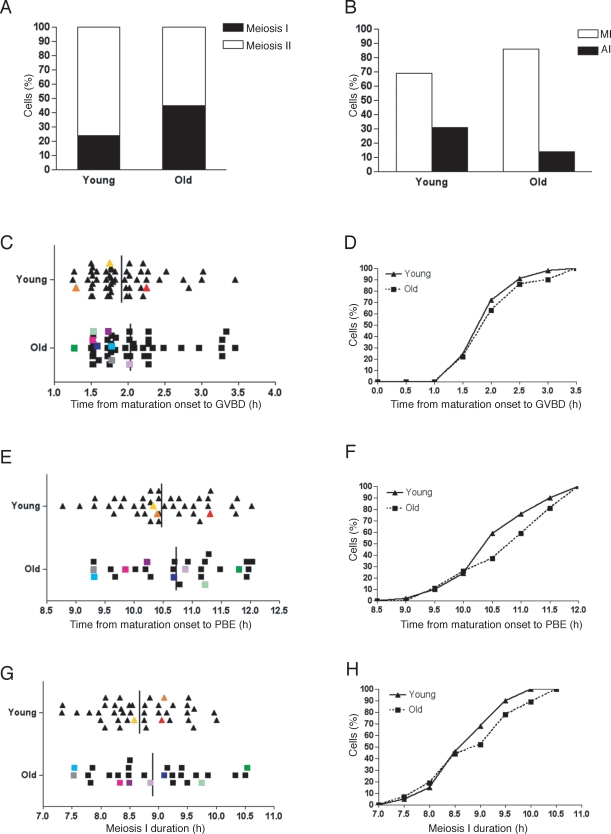
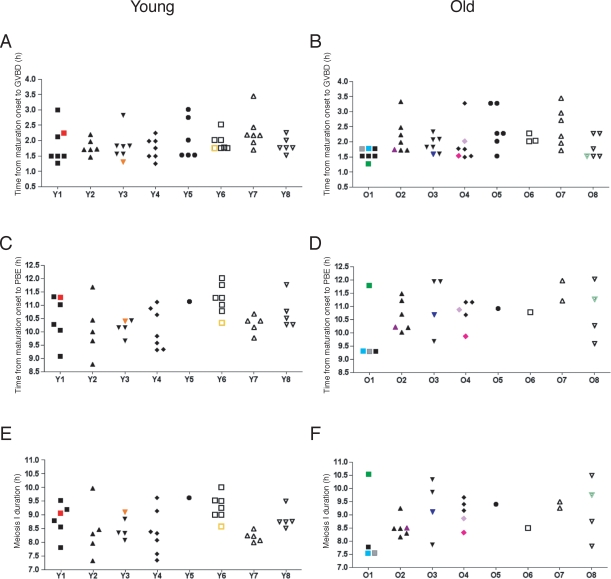
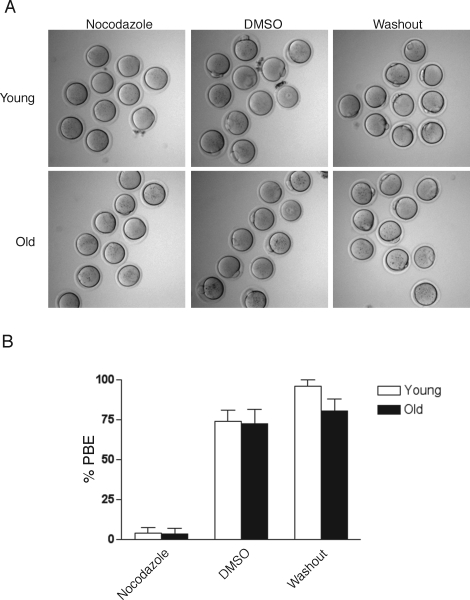
Advanced maternal age is unequivocally associated with increased aneuploidy in human eggs and infertility, but the molecular basis for this phenomenon is unknown. An age-dependent deterioration of the spindle assembly checkpoint (SAC) has been proposed as a probable cause of aneuploidy. Accurate chromosome segregation depends on correct chromosome attachment to spindle microtubules, and the SAC provides time for this process by delaying anaphase onset until all chromosomes are stably attached. If SAC function decreases with age, oocytes from reproductively old mice would enter anaphase of meiosis I (AI) prematurely, leading to chromosome segregation errors and aneuploid eggs. Although intuitively appealing, this hypothesis is largely untested. We used a natural reproductive aging mouse model to determine if a defective SAC is the primary cause of aneuploidy in eggs. We tracked the progress of individual oocytes from young and old mice through meiosis I by time-lapse microscopy and counted chromosomes in the resulting eggs. This data set allowed us to correlate the timing of AI onset with aneuploidy in individual oocytes. We found that oocytes from old mice do not enter AI prematurely compared to young counterparts despite a 4-fold increase in the incidence of aneuploidy. Moreover, we did not observe a correlation between the timing of AI onset and aneuploidy in individual oocytes. When SAC function was challenged with a low concentration of the spindle toxin nocodazole, oocytes from both young and old mice arrested at meiosis I, which is indicative of a functional checkpoint. These findings indicate that a defective SAC is unlikely the primary cause of aneuploidy associated with maternal age.
Figures
References
-
- Pellestor F, Anahory T, Hamamah S.Effect of maternal age on the frequency of cytogenetic abnormalities in human oocytes. Cytogenet Genome Res 2005; 111: 206–212.. - PubMed
-
- Hassold T, Chiu D.Maternal age-specific rates of numerical chromosome abnormalities with special reference to trisomy. Hum Genet 1985; 70: 11–17.. - PubMed
-
- Warren W, Gorringe K.A molecular model for sporadic human aneuploidy. Trends Genet 2006; 22: 218–224.. - PubMed
-
- Jones KT.Meiosis in oocytes: predisposition to aneuploidy and its increased incidence with age. Hum Reprod Update 2008; 14: 143–158.. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
