

doi: 10.1074/jbc.M109.016071.
Epub 2009 Jun 24.
Loss of NF-kappaB control and repression of Prdx6 gene transcription by reactive oxygen species-driven SMAD3-mediated transforming growth factor beta signaling
Affiliations
- PMID: 19553668
- PMCID: PMC2755684
- DOI: 10.1074/jbc.M109.016071
Item in Clipboard
Loss of NF-kappaB control and repression of Prdx6 gene transcription by reactive oxygen species-driven SMAD3-mediated transforming growth factor beta signaling
J Biol Chem.
.
No abstract available
Figures
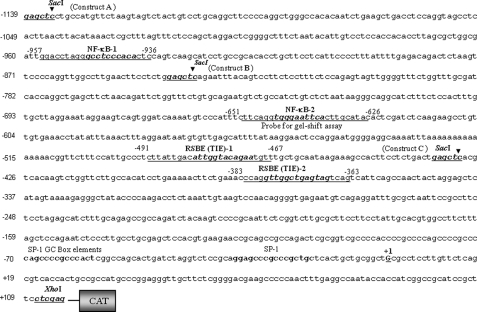
5′-Proximal regulatory region of Prdx6 gene promoter linking to CAT. Nucleotide sequences spanning from −1139 to +109 were linked to CAT reporter vector. The consensus sequences for the predicted two NF-κB (NF-κB-1 and NF-κB-2) and two RSBE/TIE binding sites are shown in boldface and italic type, respectively, and were mutated in experiments assessing the contribution of each site to promoter activity. Underlining indicates the position of oligonucleotides employed in gel shift and supershift assays. The transcription start site is indicated by +1, and SacI and XhoI restriction sites used for marking deletion mutants of Prdx6-CAT-constructs are shown in boldface type.
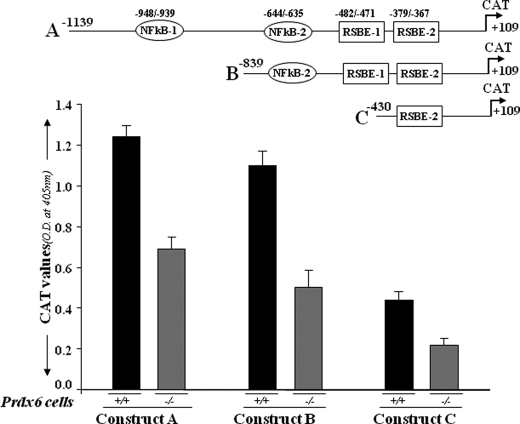
Transcriptional repression of different deletion mutants of Prdx6 gene promoter linked to CAT in Prdx6+/+ and Prdx6−/− cells. The top drawing illustrates the putative transcription-binding elements in the Prdx6 promoter. Construct A consists of two NF-κB sites (NF-κB-1 and NF-κB-2) and two repressive Smad3-binding elements (RSBE-1 and RSBE-2). Two deletion mutants were generated: Construct B with one NF-κB and two RSBE sites and Construct C with one RSBE site only. Cells were transiently transfected with Prdx6-CAT Construct A, B, or C. After 72 h, protein was extracted, and CAT activity was measured. The transfection efficiencies were normalized using a plasmid secreted alkaline phosphatase basic vector. The data represent the mean ± S.D. from three independent experiments.
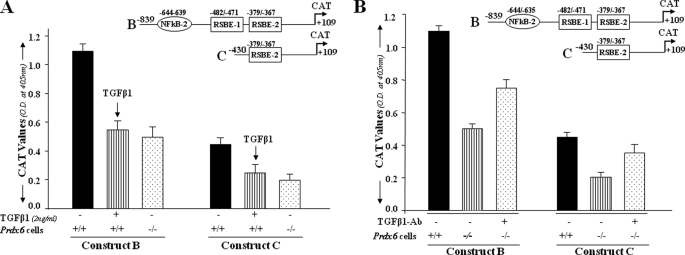
A, TGFβ1-induced repression of Prdx6 gene transcription in Prdx6+/+ cells as seen in Prdx6−/−. The cells were transiently transfected with Prdx6-CAT Construct B or C containing an RSBE site(s) as in Fig. 2. Prdx6+/+ cells (striped bars) were treated with TGFβ1 at a concentration of 2 ng/ml for 3 days, and Prdx6−/− cells and untreated Prdx6+/+ cells served as control. CAT activity of these constructs in Prdx6−/− or TGFβ1-treated or -untreated Prdx6+/+ cells was compared. Results are mean ± S.D. from three experiments. B, TGFβ-neutralizing antibody reduced the repression of Prdx6 transcription in Prdx6−/− cells. Cells were transiently transfected with either Construct B or C, and TGFβ-neutralizing antibody (R&D Systems) was added at a concentration of 5 μg/ml for 3 days followed by CAT activity determination. Striped bars indicate untreated control, and dotted bars show cells treated with TGFβ1-neutralizing antibody.
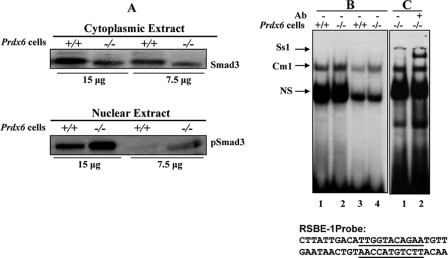
Expression levels of Smad3 and pSmad3 in the cytoplasmic and nuclear extract of Prdx6−/− cells. A, cytoplasmic and nuclear extracts were prepared and resolved by SDS-PAGE, followed by Western analysis for Smad3 and pSmad3 (compare lanes +/+ versus lanes −/−). B and C, representative gel shift and supershift assays showing binding of Smad3 present in the nuclear extracts of Prdx6−/− to RSBE-1 probe derived from the Prdx6 promoter. Nuclear extract (5 or 2 μg) from Prdx6−/− (B, lanes 2 and 4) or Prdx6+/+ (lanes 1 and 3) cells was incubated with radiolabeled probe having an SRBE-binding element (RSBE-1). Supershift assay was performed (C, lane 2) following incubation with Smad3-specific antibody (sc-6202; Santa Cruz Biotechnology). Similar results were obtained with SRBE-2 (data not shown). NS denotes nonspecific binding and demonstrates equal loading of samples.
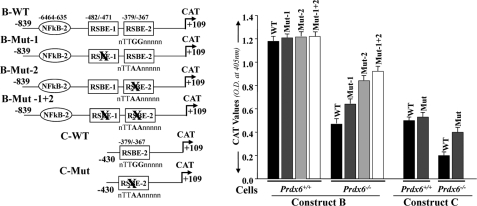
Point mutation at RSBE site(s) of the Prdx6 promoter abrogated repression of Prdx6 transcription in Prdx6−/− cells. Left, schematic illustration of wild type (Constructs B and C having RSBE/TIE) and mutated (RSBE/TIE disrupted using site-directed mutagenesis) Prdx6 promoter constructs. Right, CAT activity of engineered wild type (black bars) and mutant (gray, light gray, and open bars) promoter constructs of Prdx6 was assessed for their transcriptional activity in Prdx6−/− and Prdx6+/+ cells. All data were presented as the mean ± S.D. from at least three independent experiments.
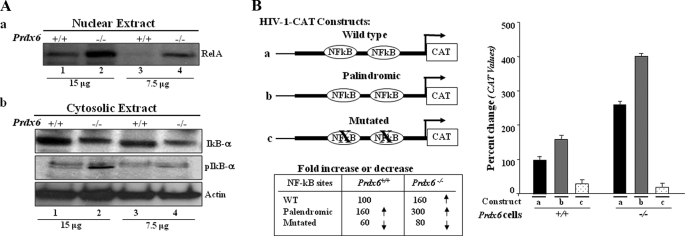
A, presence of RelA/p65 in the nuclear extract and phosphorylated/degraded form of IκBα in the cytosolic extracts of Prdx6−/− cells. a, representative Western blot displaying predominant presence of NF-κB/p65 in nuclear extracts from Prdx6−/− cells. Nuclear extracts were isolated from Prdx6−/− and Prdx6+/+ cells. Western analysis was conducted with anti-RelA/p65 (sc-7151; Santa Cruz Biotechnology) using 15 and 7.5 μg of nuclear extract (a, lanes 1 and 2). b, cytoplasmic extracts were separated, and Western analysis was conducted using pIκBα (middle, lanes 2 and 4) or IκBα (top, lanes 2 and 4) in cytosolic extracts of Prdx6−/− and Prdx6+/+ cells. B, transactivation of HIV-1LTR disclosed the presence of NF-κB signaling in Prdx6−/−-depleted cells. Relative CAT activity was measured in Prdx6−/− as well as in Prdx6+/+ cells using HIV-1LTR promoter constructs (left). Prdx6−/− and Prdx6+/+ cells were transiently transfected with either wild type pLTR-CAT (a) or pLTR-CAT-EcoR1 (b; where NF-κB sites are palindromic) or pLTR-CAT-Pst1 (c; where NF-κB sites are mutated). Promoter activity was monitored using CAT-ELISA.
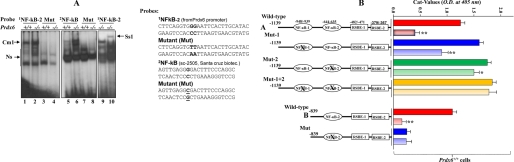
A, nuclear extract from Prdx6−/− cells strongly bound to the NF-κB-responsive element present in the Prdx6 promoter. The 32P-labeled oligonucleotide probes 1NF-κB-2 and 2NF-κB (as standard control) and their mutants were incubated with nuclear extract isolated from Prdx6−/− and Prdx6+/+ cells, and a gel shift assay was performed. A supershift assay using Rel/P65 antibody following incubation with Prdx6−/− nuclear protein extract was carried out (Ss1; lane 10). B, point mutation at NF-κB sites identified by gel shift assay experiment (NF-κB-2) in the Prdx6 promoter Construct B abolished its transcriptional activity, whereas disruption of NF-κB-1 and/or NF-κB-2 in Construct A increased the promoter activity. Prdx6+/+ or Prdx6−/− cells were transiently transfected with equal amounts of wild type and their mutant promoter-driven CAT reporters (left panel, showing schematic representation of constructs in the experiment), and the CAT activity was measured. For Construct A, CAT activity of promoter mutated at NF-κB sites (Mut-1 (blue bar), Mut-2 (green bar), or Mut1 + Mut-2 (yellow bar)) was compared with the activity of wild type promoter constructs (WT (red bar)). In the same set of experiments, cells were also cotransfected with dominant-negative Iκ-Bα mutant (Iκ-BαAA; light red bar, light blue bar, and light yellow bar). For Construct B, cells were transfected either with wild-type NF-κB or its mutant, and CAT activity was monitored (B, WT versus Mut). In another set of experiments, cells were cotransfected with Iκ-BαAA (Construct B, light red and light blue bars). The CAT vector showed insignificant activity (data not shown).

A, PRDX6 delivery to Prdx6−/− cells restored the Prdx6 promoter activity by attenuating Smad3 binding to RSBE. Prdx6−/− and Prdx6+/+ cells were cultured in the presence (4 μg/ml) or absence of PRDX6 (4). Nuclear extract was isolated and incubated with radiolabeled RSBE-1 probe, and a gel shift assay was performed. Binding affinity of nuclear extract from Prdx6−/− cells supplied with recombinant PRDX6 (lane 3, Cm1; see Figs. 1 and 4 for RSBE-1) was compared with nuclear extract of Prdx6−/− cells (lane 2) to radiolabeled oligonucleotide probe containing RSBE. B, gel shift assay showing binding of nuclear extract from Prdx6−/− cells to radiolabeled probe containing NF-κB binding site or its mutant probe (NF-κB-2; see Fig. 7); binding activity of nuclear extracts isolated from cells treated with PRDX6 (lanes 2 and 3) compared with that of cells where TAT-HA-PRDX6 was not added (lane 1). C, Prdx6−/− cells showing higher expression of ROS. An H2-DCF-DA assay was conducted to monitor the effect of NAC on levels of ROS (C, gray bars). D and E, cells were transiently transfected with different Prdx6-CAT constructs and were cultured with or without PRDX6 for 3 consecutive days (4 μg/ml) or treated with NAC (1 mm ), an antioxidant. After 72 h, cells were harvested, and CAT activity was monitored (D, gray bars) or NAC (E, gray bars).
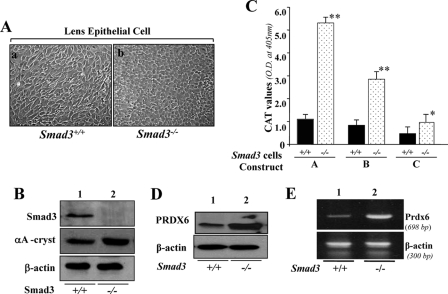
A, photomicrograph of Smad3+/+ and Smad3−/− lens epithelial cells cultured in vitro and validation of their integrity. Smad3+/+ (a) and Smad3−/− (b) LECs were isolated from their corresponding mouse lenses and maintained in DMEM plus 10% fetal bovine serum (1). B, integrity of these cells was validated using αA-crystallin antibody (middle), a specific marker of LECs, and Smad3-specific antibody (top) using Western analysis. Bottom, β-actin, an internal control. C–E, unlike Prdx6−/− cells, Smad3−/− cells displayed enhanced Prdx6 promoter activity and increased expression of PRDX6 protein. C, a representative of transactivation assay experiments showing CAT activities in Smad3−/− and Smad3+/+ cells cultured in serum-depleted media. Cells were transfected with equal amounts of Prdx6 promoter linked to CAT. CAT activity was measured as described under “Experimental Procedures.” CAT activity of deletion mutant constructs in Smad3−/− cells (dotted bar) and Smad3+/+ cells (black bars) was monitored. Western analysis (D) and RT-PCR (E) showing expression of PRDX6 protein and mRNA in Smad3−/− cells, respectively.
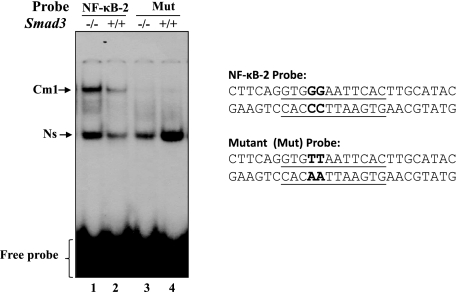
NF-κB in the nuclear extract from Smad3−/− cells was transcriptionally active, bound strongly to its responsive element(s). A gel shift assay was conducted with the nuclear extracts isolated from Smad3−/− and Smad3+/+ cells cultured in serum-depleted medium as described earlier following incubation with radiolabeled oligonucleotide probe, NF-κB-2 (lane 1), or its mutant. Similar results were obtained with NF-κB-1 probe (data not shown).

A, Smad3−/− cells engendered resistance against TGFβ-induced insults. Photomicrographs of Smad3−/− (left) or Smad3+/+ (right) cells treated with TGFβ1 (2 ng/ml). Cells were cultured for 24 h in DMEM containing 10% fetal bovine serum and then washed. The medium was replaced with 0.2% bovine serum albumin in DMEM with or without TGFβ1 for 24 or 48 h. The arrows indicate dead cells. B, cell viability of Smad3−/− cells against TGFβ1-induced insults was estimated using an MTS assay (black bar). C, TGFβ1 failed to suppress Prdx6 expression in Smad3−/− cells (top, right lane). Western analysis was conducted using cellular extract isolated from Smad3−/− and Smad3+/+ cells treated or untreated with TGFβ1. Cell lysate was prepared using radioimmunoprecipitation buffer, samples were resolved on SDS-PAGE, and Western analysis was done with anti-PRDX6 antibody. β-actin was used as loading control. D, Smad3−/− LECs showed resistance against H2O2-induced oxidative stress-mediated damage. Smad3+/+ (black bars) and Smad3−/− (gray bar) cells were exposed to H2O2 at 100 or 200 μm for 2 h. Cell viability was estimated after 24 h of recovery using an MTS assay.
Similar articles
-
Curcumin abates hypoxia-induced oxidative stress based-ER stress-mediated cell death in mouse hippocampal cells (HT22) by controlling Prdx6 and NF-κB regulation.Am J Physiol Cell Physiol. 2013 Apr 1;304(7):C636-55. doi: 10.1152/ajpcell.00345.2012. Epub 2013 Jan 30. Am J Physiol Cell Physiol. 2013. PMID: 23364261 Free PMC article.
-
Peroxiredoxin-6 Negatively Regulates Bactericidal Activity and NF-κB Activity by Interrupting TRAF6-ECSIT Complex.Front Cell Infect Microbiol. 2017 Mar 24;7:94. doi: 10.3389/fcimb.2017.00094. eCollection 2017. Front Cell Infect Microbiol. 2017. PMID: 28393051 Free PMC article.
-
Peroxiredoxin 6 expression is inversely correlated with nuclear factor-κB activation during Clonorchis sinensis infestation.Free Radic Biol Med. 2016 Oct;99:273-285. doi: 10.1016/j.freeradbiomed.2016.08.016. Epub 2016 Aug 20. Free Radic Biol Med. 2016. PMID: 27554973
-
Delivery of a protein transduction domain-mediated Prdx6 protein ameliorates oxidative stress-induced injury in human and mouse neuronal cells.Am J Physiol Cell Physiol. 2016 Jan 1;310(1):C1-16. doi: 10.1152/ajpcell.00229.2015. Epub 2015 Oct 7. Am J Physiol Cell Physiol. 2016. PMID: 26447207 Free PMC article.
-
Runx2/Smad3 complex negatively regulates TGF-β-induced connective tissue growth factor gene expression in vascular smooth muscle cells.J Atheroscler Thromb. 2012;19(1):23-35. doi: 10.5551/jat.9753. Epub 2011 Oct 8. J Atheroscler Thromb. 2012. PMID: 21986102
Cited by
-
Curcumin abates hypoxia-induced oxidative stress based-ER stress-mediated cell death in mouse hippocampal cells (HT22) by controlling Prdx6 and NF-κB regulation.Am J Physiol Cell Physiol. 2013 Apr 1;304(7):C636-55. doi: 10.1152/ajpcell.00345.2012. Epub 2013 Jan 30. Am J Physiol Cell Physiol. 2013. PMID: 23364261 Free PMC article.
-
Engineered Sumoylation-Deficient Prdx6 Mutant Protein-Loaded Nanoparticles Provide Increased Cellular Defense and Prevent Lens Opacity.Antioxidants (Basel). 2021 Aug 4;10(8):1245. doi: 10.3390/antiox10081245. Antioxidants (Basel). 2021. PMID: 34439493 Free PMC article.
-
Vitamin D prevents hypoxia/reoxygenation-induced blood-brain barrier disruption via vitamin D receptor-mediated NF-kB signaling pathways.PLoS One. 2015 Mar 27;10(3):e0122821. doi: 10.1371/journal.pone.0122821. eCollection 2015. PLoS One. 2015. PMID: 25815722 Free PMC article.
-
Chlorella vulgaris modulates the expression of senescence-associated genes in replicative senescence of human diploid fibroblasts.Mol Biol Rep. 2020 Jan;47(1):369-379. doi: 10.1007/s11033-019-05140-8. Epub 2019 Oct 22. Mol Biol Rep. 2020. PMID: 31642042
-
Therapeutic prospects and potential mechanisms of Prdx6: as a novel target in musculoskeletal disorders.Front Physiol. 2025 Apr 17;16:1524100. doi: 10.3389/fphys.2025.1524100. eCollection 2025. Front Physiol. 2025. PMID: 40313876 Free PMC article. Review.
References
-
- Fatma N., Kubo E., Sharma P., Beier D. R., Singh D. P. (2005) Cell Death Differ. 12,734–750 - PubMed
-
- Fatma N., Singh D. P., Shinohara T., Chylack L. T., Jr. (2001) J. Biol. Chem. 276,48899–48907 - PubMed
-
- Fisher A. B., Dodia C., Manevich Y., Chen J. W., Feinstein S. I. (1999) J. Biol. Chem. 274,21326–21334 - PubMed
-
- Kubo E., Fatma N., Akagi Y., Beier D. R., Singh S. P., Singh D. P. (2008) Am. J. Physiol. Cell Physiol. 294,C842–C855 - PubMed
-
- Kubo E., Miyazawa T., Fatma N., Akagi Y., Singh D. P. (2006) Mech. Ageing Dev. 127,249–256 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
