

Odor representations in olfactory cortex: "sparse" coding, global inhibition, and oscillations
- PMID: 19555653
- PMCID: PMC2702531
- DOI: 10.1016/j.neuron.2009.05.022
Odor representations in olfactory cortex: "sparse" coding, global inhibition, and oscillations
Abstract
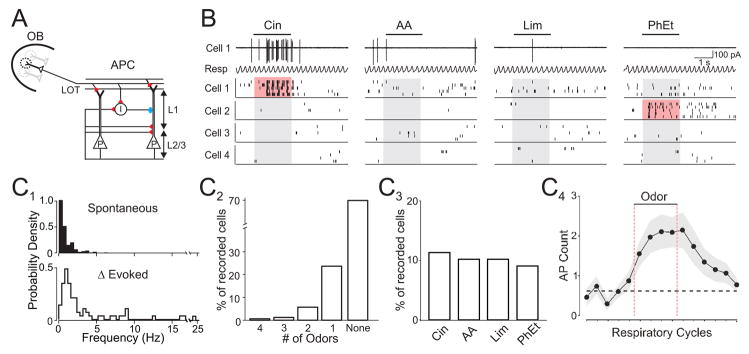
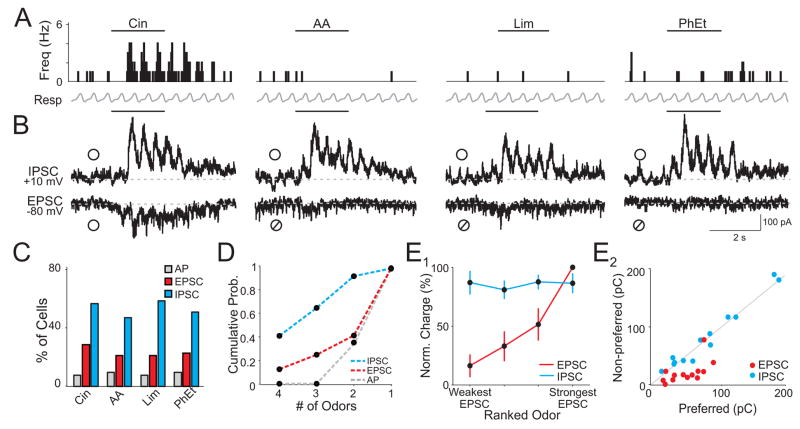
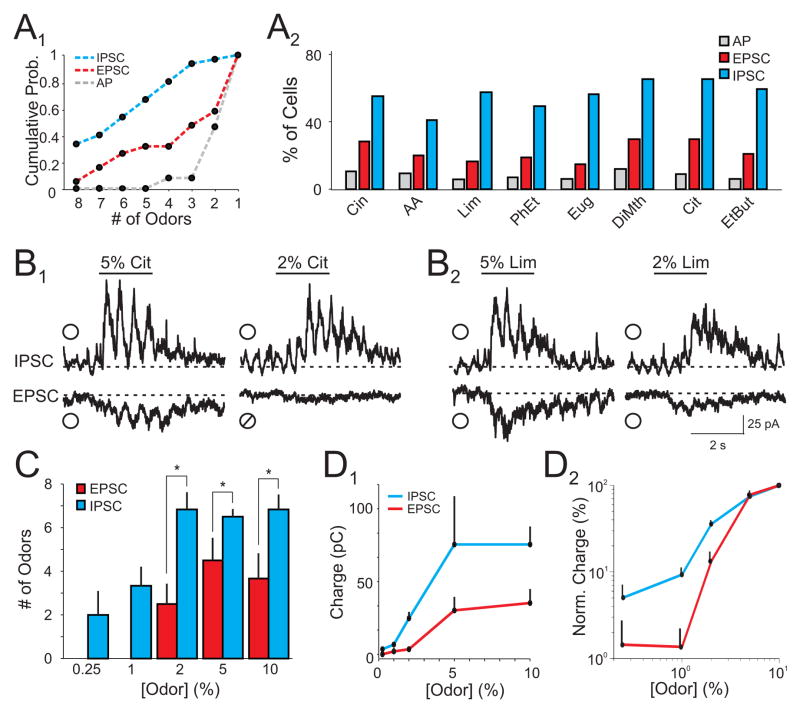
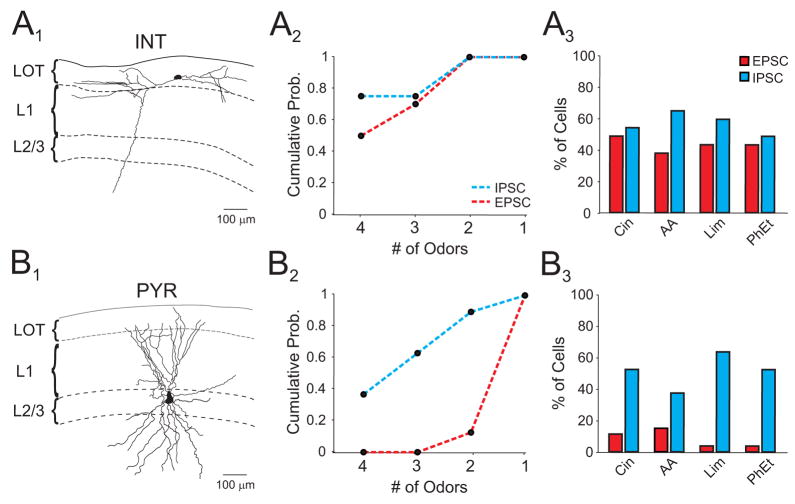
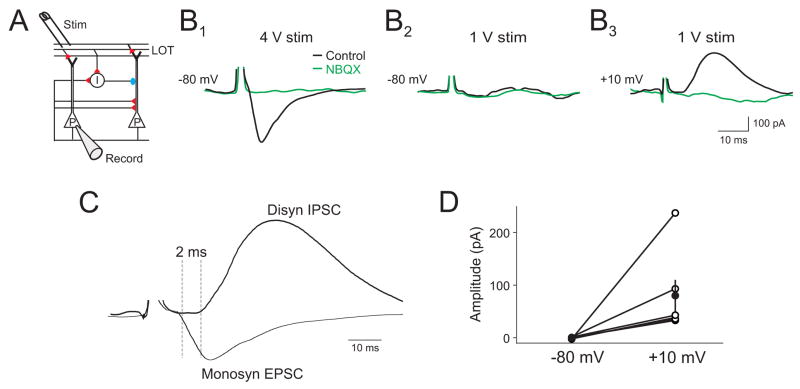
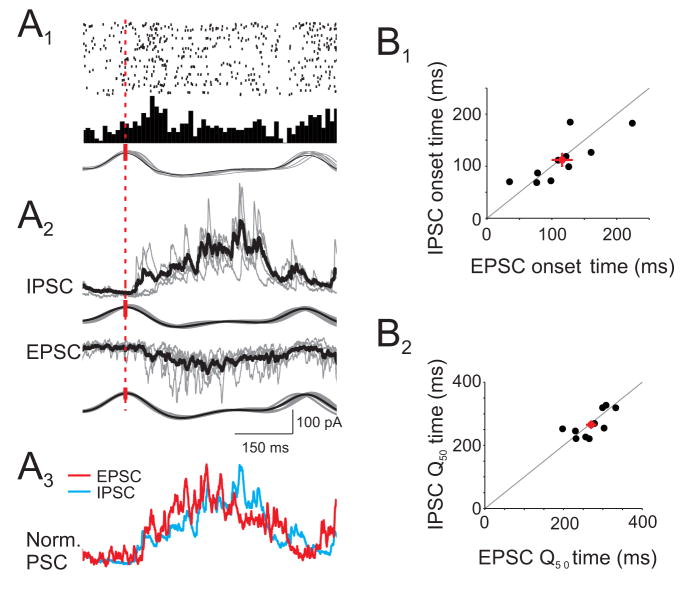
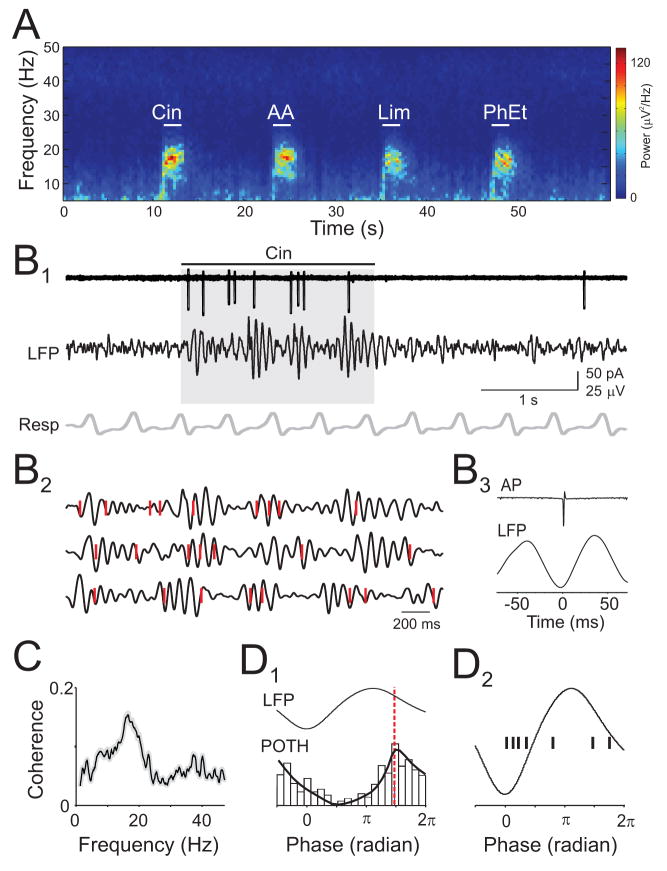
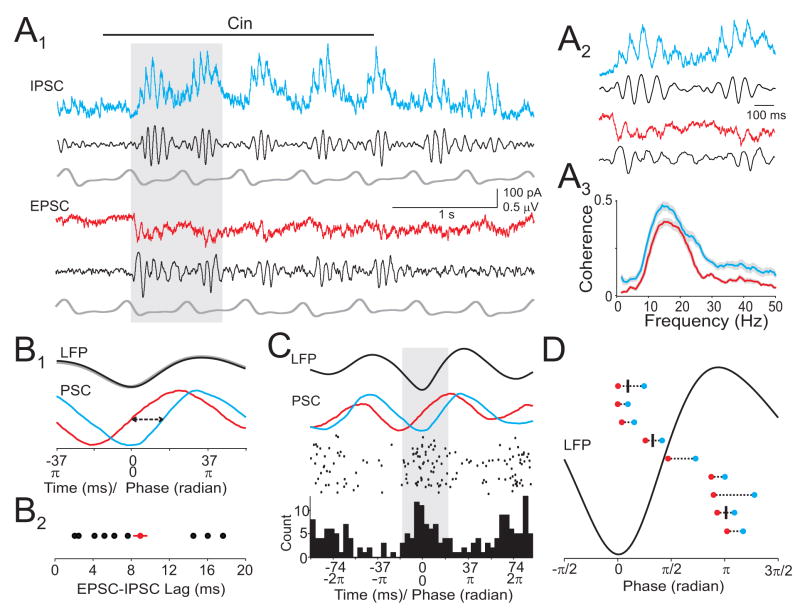
The properties of cortical circuits underlying central representations of sensory stimuli are poorly understood. Here we use in vivo cell-attached and whole-cell voltage-clamp recordings to reveal how excitatory and inhibitory synaptic input govern odor representations in rat primary olfactory (piriform) cortex. We show that odors evoke spiking activity that is sparse across the cortical population. We find that unbalanced synaptic excitation and inhibition underlie sparse activity: inhibition is widespread and broadly tuned, while excitation is less common and odor-specific. "Global" inhibition can be explained by local interneurons that receive ubiquitous and nonselective odor-evoked excitation. In the temporal domain, while respiration imposes a slow rhythm to olfactory cortical responses, odors evoke fast (15-30 Hz) oscillations in synaptic activity. Oscillatory excitation precedes inhibition, generating brief time windows for precise and temporally sparse spike output. Together, our results reveal that global inhibition and oscillations are major synaptic mechanisms shaping odor representations in olfactory cortex.
Figures
Comment in
-
Inhibition acts globally to shape olfactory cortical tuning.Neuron. 2009 Jun 25;62(6):750-2. doi: 10.1016/j.neuron.2009.06.004. Neuron. 2009. PMID: 19555643
References
-
- Anderson JS, Carandini M, Ferster D. Orientation tuning of input conductance, excitation, and inhibition in cat primary visual cortex. J Neurophysiol. 2000;84:909–926. - PubMed
-
- Bathellier B, Buhl DL, Accolla R, Carleton A. Dynamic ensemble odor coding in the mammalian olfactory bulb: sensory information at different timescales. Neuron. 2008;57:586–598. - PubMed
-
- Buonviso N, Amat C, Litaudon P. Respiratory modulation of olfactory neurons in the rodent brain. Chem Senses. 2006;31:145–154. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
