

Dissecting the essentiality of the bifunctional trypanothione synthetase-amidase in Trypanosoma brucei using chemical and genetic methods
- PMID: 19558432
- PMCID: PMC2784880
- DOI: 10.1111/j.1365-2958.2009.06761.x
Dissecting the essentiality of the bifunctional trypanothione synthetase-amidase in Trypanosoma brucei using chemical and genetic methods
Abstract
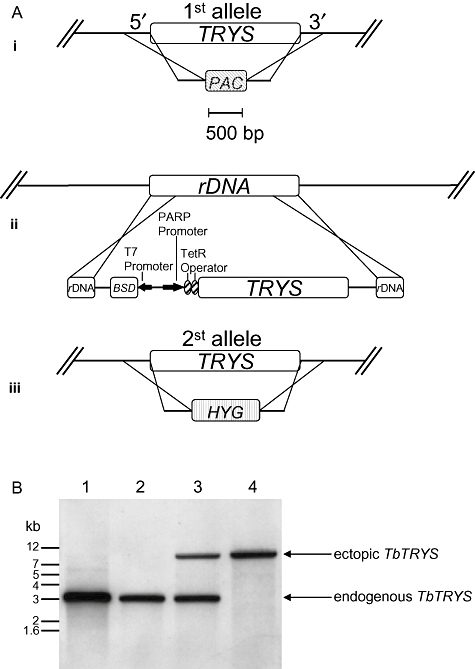
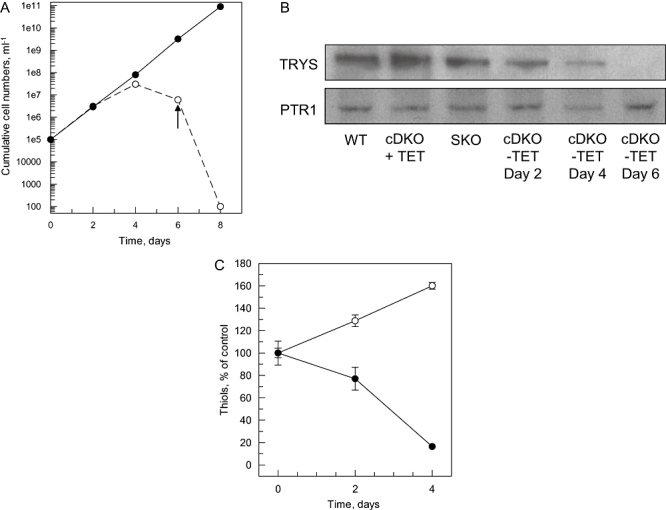
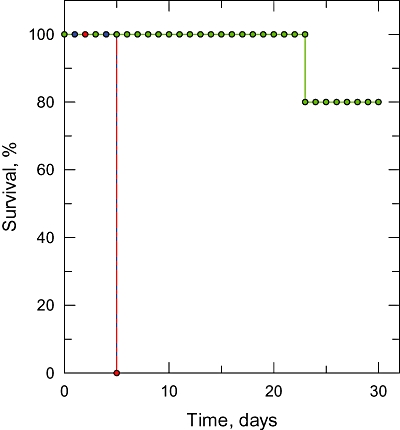
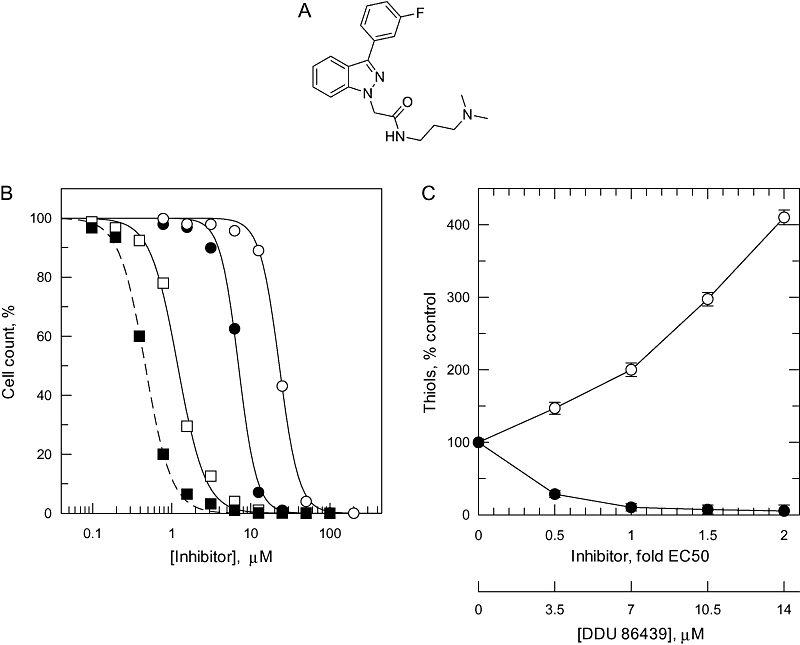
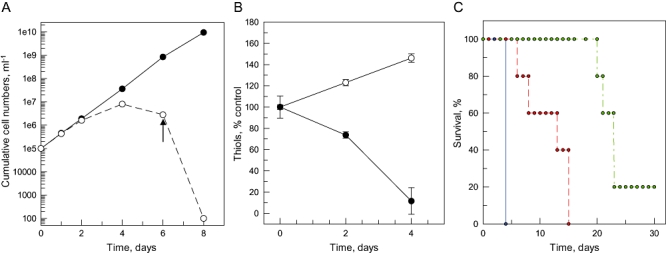
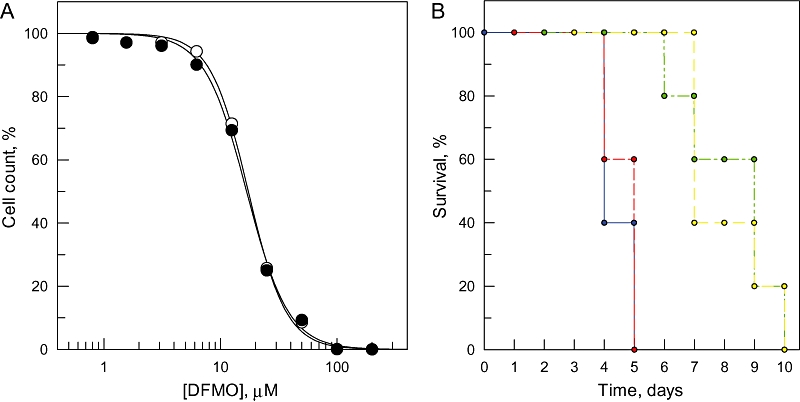
The bifunctional trypanothione synthetase-amidase (TRYS) comprises two structurally distinct catalytic domains for synthesis and hydrolysis of trypanothione (N(1),N(8)-bis(glutathionyl)spermidine). This unique dithiol plays a pivotal role in thiol-redox homeostasis and in defence against chemical and oxidative stress in trypanosomatids. A tetracycline-dependent conditional double knockout of TRYS (cDKO) was generated in bloodstream Trypanosoma brucei. Culture of cDKO parasites without tetracycline induction resulted in loss of trypanothione and accumulation of glutathione, followed by growth inhibition and cell lysis after 6 days. In the absence of inducer, cDKO cells were unable to infect mice, confirming that this enzyme is essential for virulence in vivo as well as in vitro. To establish whether both enzymatic functions were essential, an amidase-dead mutant cDKO line was generated. In the presence of inducer, this line showed decreased growth in vitro and decreased virulence in vivo, indicating that the amidase function is not absolutely required for viability. The druggability of TRYS was assessed using a potent small molecule inhibitor developed in our laboratory. Growth inhibition correlated in rank order cDKO, single KO, wild-type and overexpressing lines and produced the predicted biochemical phenotype. The synthetase function of TRYS is thus unequivocally validated as a drug target by both chemical and genetic methods.
Figures
References
-
- Alibu VP, Richter C, Voncken F, Marti G, Shahi S, Renggli CK, et al. The role of Trypanosoma brucei MRPA in melarsoprol susceptibility. Mol Biochem Parasitol. 2006;146:38–44. - PubMed
-
- Ariyanayagam MR, Fairlamb AH. Ovothiol and trypanothione as antioxidants in trypanosomatids. Mol Biochem Parasitol. 2001;115:189–198. - PubMed
-
- Bacchi CJ, Yarlett N. Polyamine metabolism. In: Marr JJ, Müller M, editors. Biochemistry and Molecular Biology of Parasites. London: Academic Press; 1995. pp. 119–131.
-
- Barrett MP, Fairlamb AH. The biochemical basis of arsenical-diamidine cross-resistance in African trypanosomes. Parasitol Today. 1999;15:136–140. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
