

Autocrine IL-10 induces hallmarks of alternative activation in macrophages and suppresses antituberculosis effector mechanisms without compromising T cell immunity
- PMID: 19561100
- PMCID: PMC2735238
- DOI: 10.4049/jimmunol.0803567
Autocrine IL-10 induces hallmarks of alternative activation in macrophages and suppresses antituberculosis effector mechanisms without compromising T cell immunity
Abstract
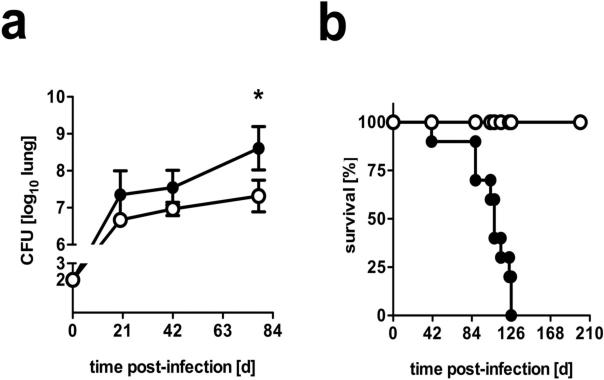
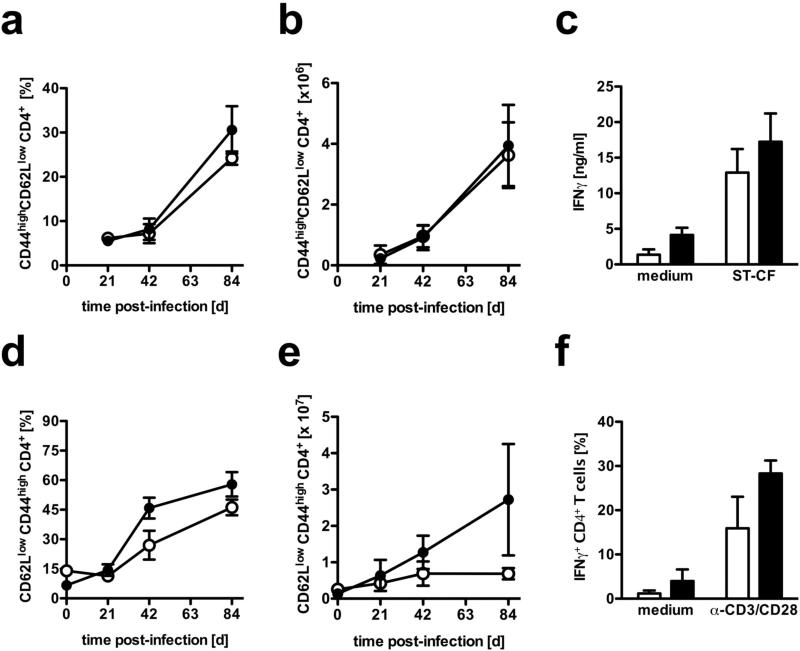
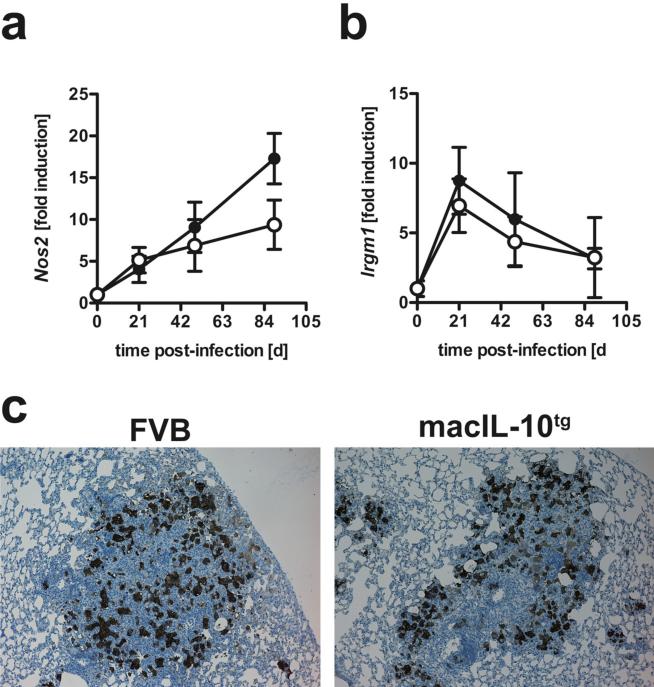
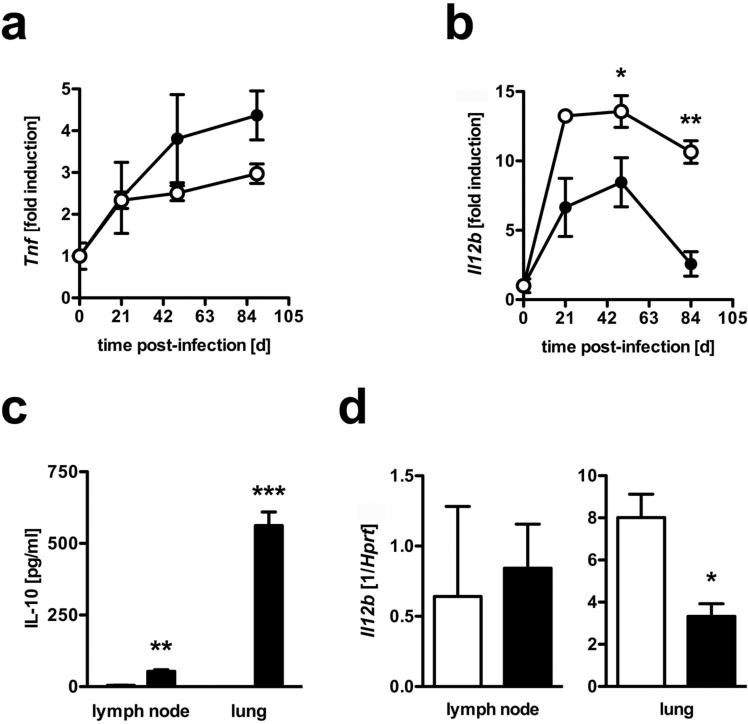
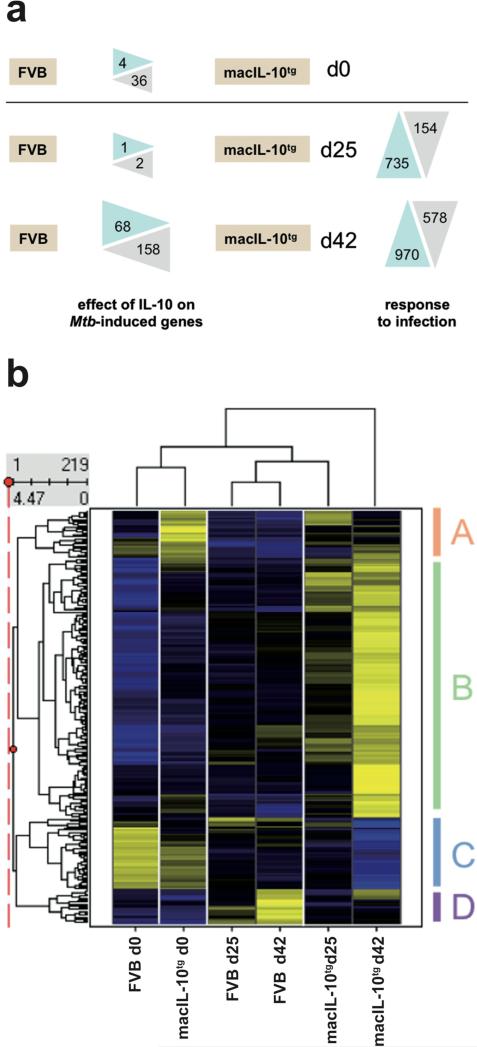
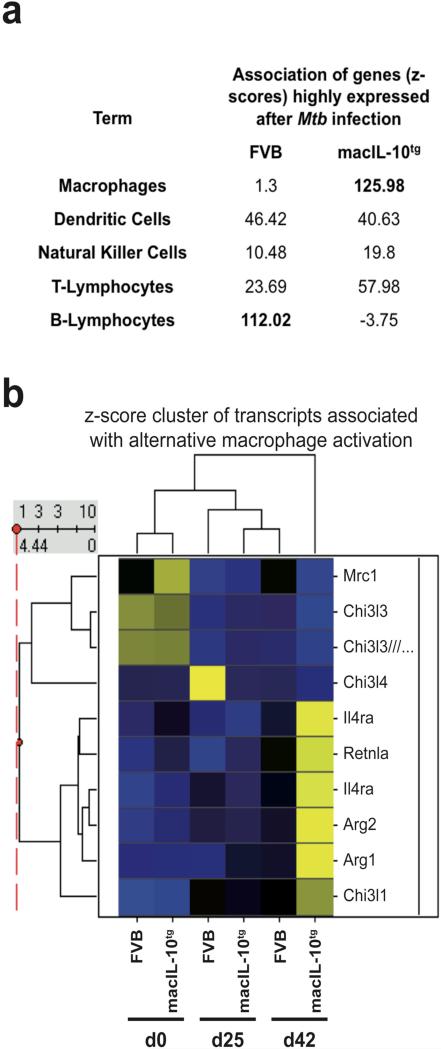
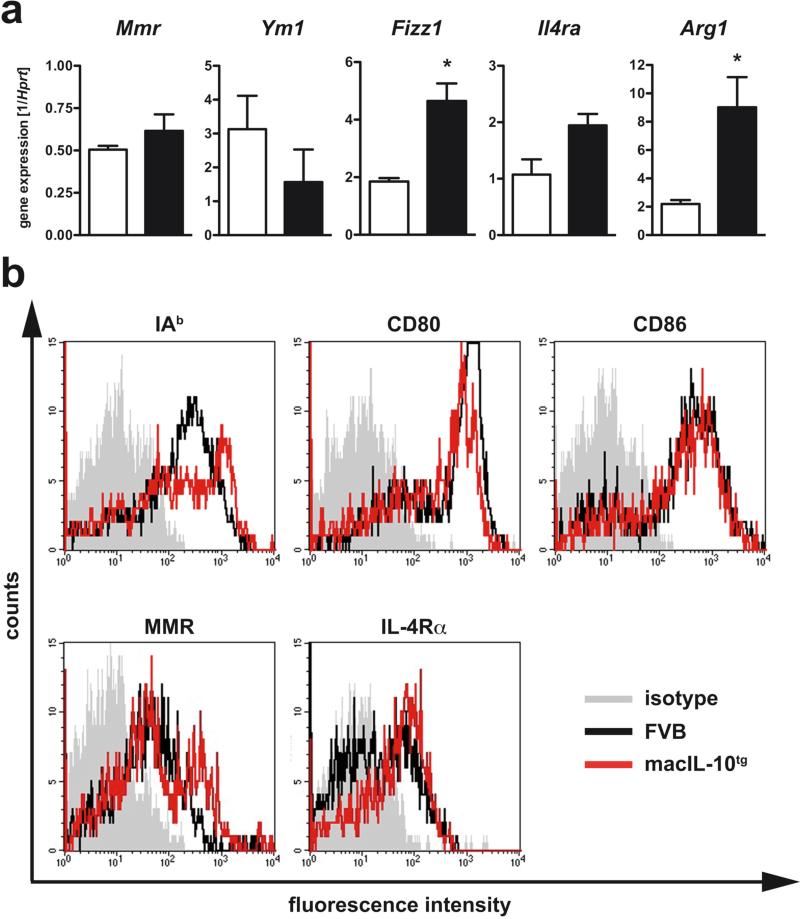
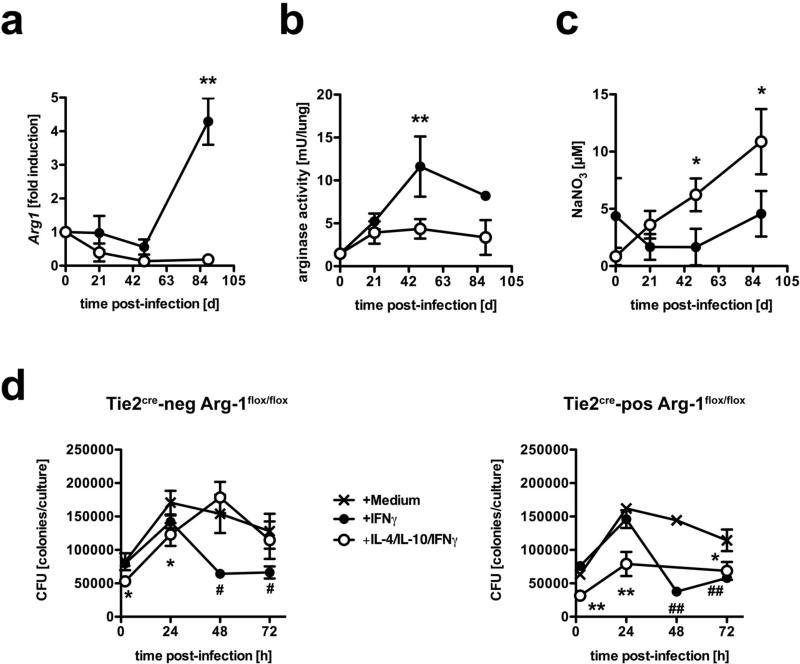
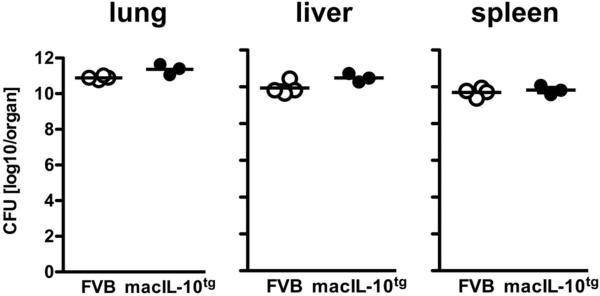
Elevated IL-10 has been implicated in reactivation tuberculosis (TB). Since macrophages rather than T cells were reported to be the major source of IL-10 in TB, we analyzed the consequences of a macrophage-specific overexpression of IL-10 in transgenic mice (macIL-10-transgenic) after aerosol infection with Mycobacterium tuberculosis (Mtb). MacIL-10 transgenic mice were more susceptible to chronic Mtb infection than nontransgenic littermates, exhibiting higher bacterial loads in the lung after 12 wk of infection and dying significantly earlier than controls. The differentiation, recruitment, and activation of Th1 cells as well as the induction of IFN-gamma-dependent effector genes against Mtb were not affected by macrophage-derived IL-10. However, microarray analysis of pulmonary gene expression revealed patterns characteristic of alternative macrophage activation that were overrepresented in Mtb-infected macIL-10 transgenic mice. Importantly, arginase-1 gene expression and activity were strikingly enhanced in transgenic mice accompanied by a reduced production of reactive nitrogen intermediates. Moreover, IL-10-dependent arginase-1 induction diminished antimycobacterial effector mechanisms in macrophages. Taken together, macrophage-derived IL-10 triggers aspects of alternative macrophage activation and promotes Mtb recrudescence independent of overt effects on anti-TB T cell immunity.
Figures
References
-
- Kaufmann SH. Tuberculosis: back on the immunologists’ agenda. Immunity. 2006;24:351–357. - PubMed
-
- Flynn JL, Goldstein MM, Chan J, Triebold KJ, Pfeffer K, Lowenstein CJ, Schreiber R, Mak TW, Bloom BR. Tumor necrosis factor-alpha is required in the protective immune response against Mycobacterium tuberculosis in mice. Immunity. 1995;2:561–572. - PubMed
-
- MacMicking JD, Taylor GA, McKinney JD. Immune control of tuberculosis by IFN-gamma-inducible LRG-47. Science. 2003;302:654–659. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Research Materials
