

Adiponectin deficiency: a model of pulmonary hypertension associated with pulmonary vascular disease
- PMID: 19561137
- PMCID: PMC2739777
- DOI: 10.1152/ajplung.90599.2008
Adiponectin deficiency: a model of pulmonary hypertension associated with pulmonary vascular disease
Abstract
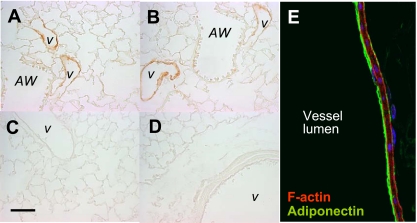
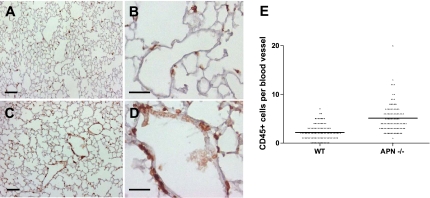
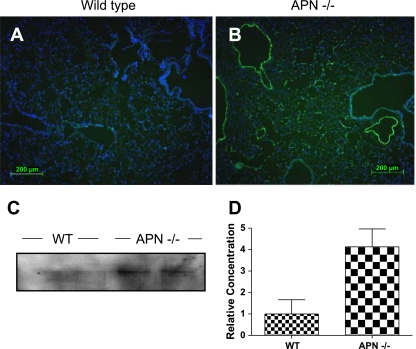
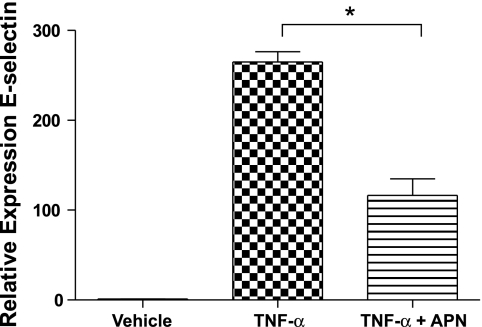
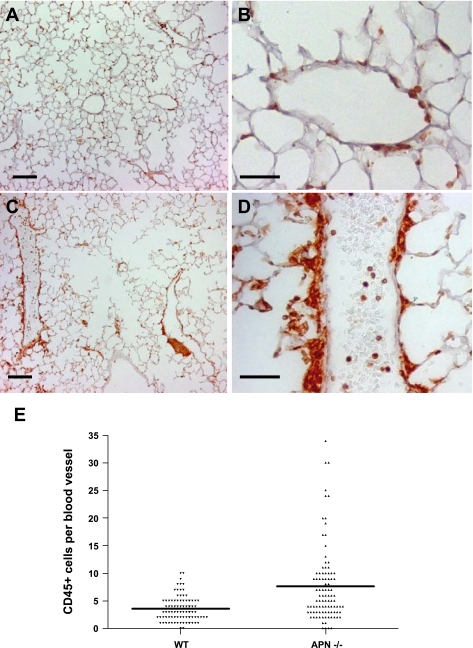
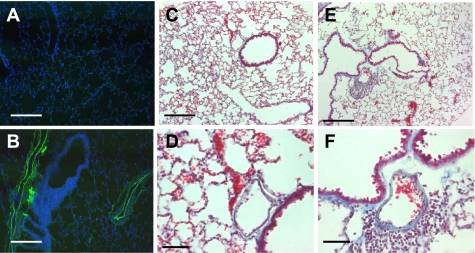
Adiponectin (APN) is an adipocyte-derived factor that exists at high concentrations in serum and has anti-inflammatory and systemic vascular-protective properties. In this study, we investigated the role of APN in pulmonary vascular homeostasis. We found that APN localizes to the luminal side of blood vessels in lung and acts in vitro to block TNF-alpha-induced E-selectin upregulation in pulmonary artery endothelial cells. Targeted deletion of the APN gene in mice leads to a vascular phenotype in lung characterized by E-selectin upregulation and age-dependent increases in perivascular inflammatory cell infiltration and pulmonary arterial pressures. Taken together, these findings demonstrate an important role for APN in lung vascular homeostasis and suggest that APN-deficient states may contribute to the pathogenesis of inflammatory pulmonary vascular disease and to the development of pulmonary hypertension.
Figures
References
-
- Arita Y, Kihara S, Ouchi N, Takahashi M, Maeda K, Miyagawa J, Hotta K, Shimomura I, Nakamura T, Miyaoka K, Kuriyama H, Nishida M, Yamashita S, Okubo K, Matsubara K, Muraguchi M, Ohmoto Y, Funahashi T, Matsuzawa Y. Paradoxical decrease of an adipose-specific protein, adiponectin, in obesity. Biochem Biophys Res Commun 257: 79–83, 1999. - PubMed
-
- Dorfmuller P, Perros F, Balabanian K, Humbert M. Inflammation in pulmonary arterial hypertension. Eur Respir J 22: 358–363, 2003. - PubMed
-
- Farber HW, Loscalzo J. Pulmonary arterial hypertension. N Engl J Med 351: 1655–1665, 2004. - PubMed
-
- Hattori Y, Nakano Y, Hattori S, Tomizawa A, Inukai K, Kasai K. High molecular weight adiponectin activates AMPK and suppresses cytokine-induced NF-kappaB activation in vascular endothelial cells. FEBS Lett 582: 1719–1724, 2008. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Miscellaneous