

ANO2 is the cilial calcium-activated chloride channel that may mediate olfactory amplification
- PMID: 19561302
- PMCID: PMC2702256
- DOI: 10.1073/pnas.0903304106
ANO2 is the cilial calcium-activated chloride channel that may mediate olfactory amplification
Abstract
For vertebrate olfactory signal transduction, a calcium-activated chloride conductance serves as a major amplification step. However, the molecular identity of the olfactory calcium-activated chloride channel (CaCC) is unknown. Here we report a proteomic screen for cilial membrane proteins of mouse olfactory sensory neurons (OSNs) that identified all the known olfactory transduction components as well as Anoctamin 2 (ANO2). Ano2 transcripts were expressed specifically in OSNs in the olfactory epithelium, and ANO2::EGFP fusion protein localized to the OSN cilia when expressed in vivo using an adenoviral vector. Patch-clamp analysis revealed that ANO2, when expressed in HEK-293 cells, forms a CaCC and exhibits channel properties closely resembling the native olfactory CaCC. Considering these findings together, we propose that ANO2 constitutes the olfactory calcium-activated chloride channel.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
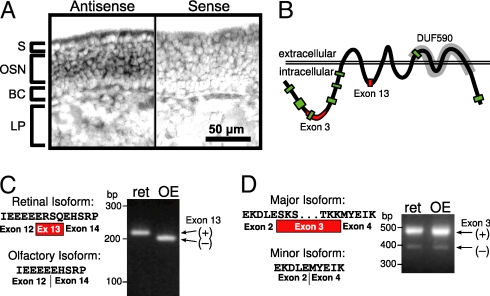
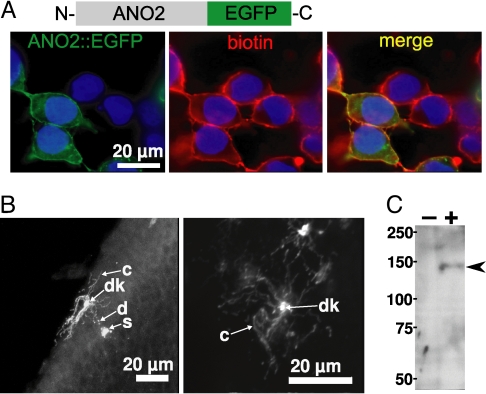
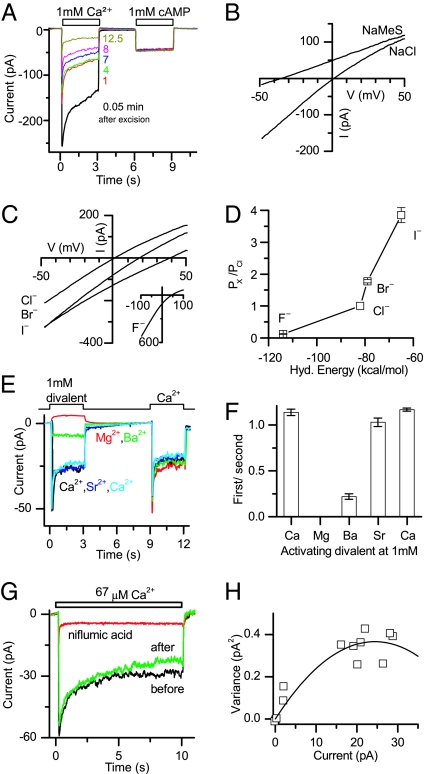
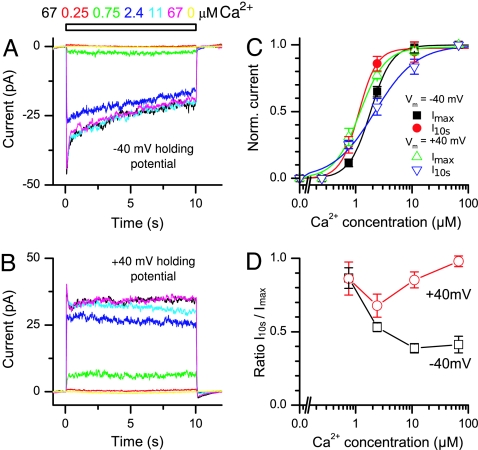
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
