

Structure of N-terminal domain of NPC1 reveals distinct subdomains for binding and transfer of cholesterol
- PMID: 19563754
- PMCID: PMC2739658
- DOI: 10.1016/j.cell.2009.03.049
Structure of N-terminal domain of NPC1 reveals distinct subdomains for binding and transfer of cholesterol
Abstract
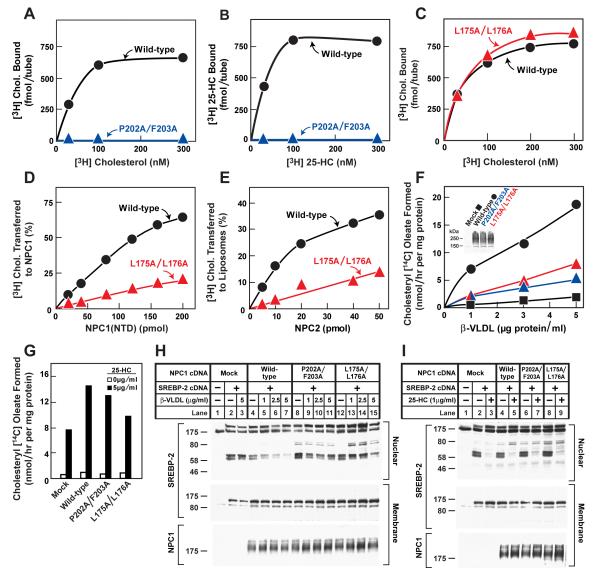
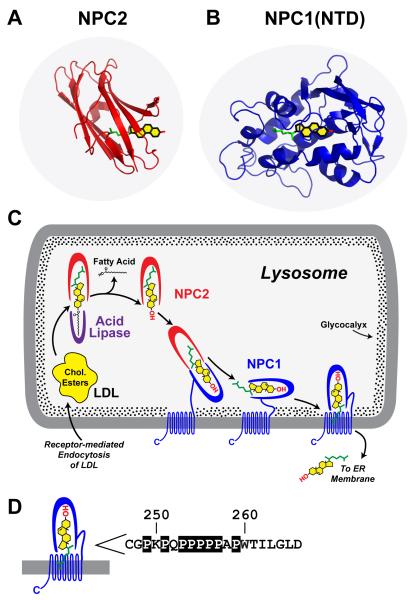
LDL delivers cholesterol to lysosomes by receptor-mediated endocytosis. Exit of cholesterol from lysosomes requires two proteins, membrane-bound Niemann-Pick C1 (NPC1) and soluble NPC2. NPC2 binds cholesterol with its isooctyl side chain buried and its 3beta-hydroxyl exposed. Here, we describe high-resolution structures of the N-terminal domain (NTD) of NPC1 and complexes with cholesterol and 25-hydroxycholesterol. NPC1(NTD) binds cholesterol in an orientation opposite to NPC2: 3beta-hydroxyl buried and isooctyl side chain exposed. Cholesterol transfer from NPC2 to NPC1(NTD) requires reorientation of a helical subdomain in NPC1(NTD), enlarging the opening for cholesterol entry. NPC1 with point mutations in this subdomain (distinct from the binding subdomain) cannot accept cholesterol from NPC2 and cannot restore cholesterol exit from lysosomes in NPC1-deficient cells. We propose a working model wherein after lysosomal hydrolysis of LDL-cholesteryl esters, cholesterol binds NPC2, which transfers it to NPC1(NTD), reversing its orientation and allowing insertion of its isooctyl side chain into the outer lysosomal membranes.
Figures
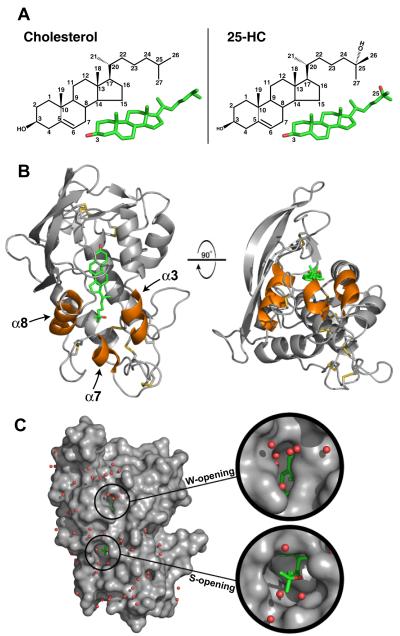
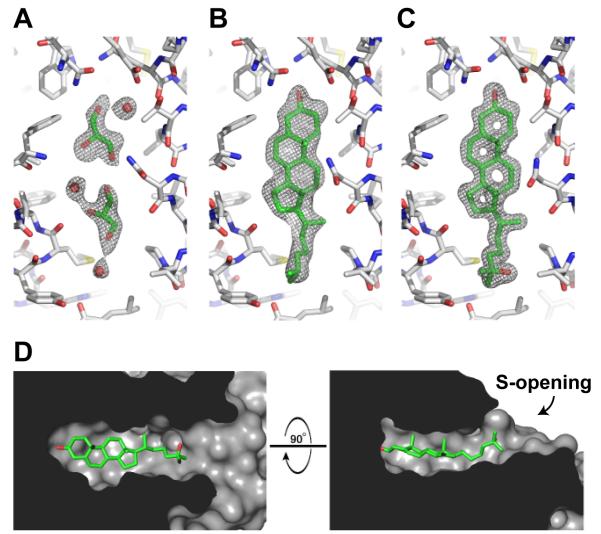
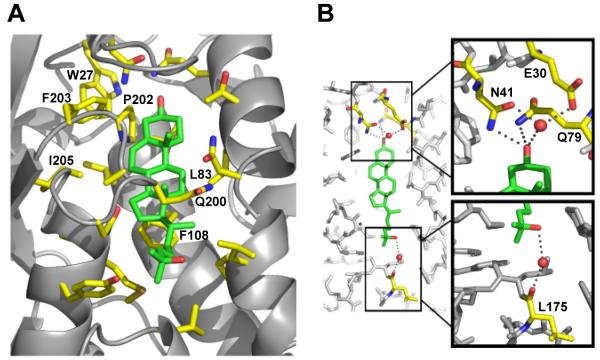
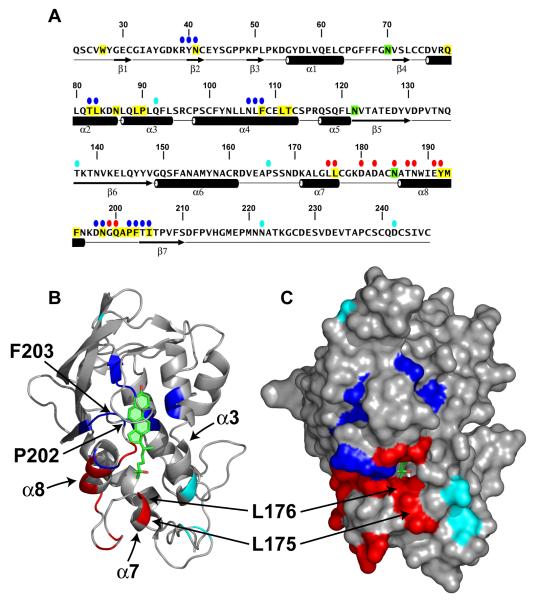
Comment in
-
Spreading the wealth: Niemann-Pick type C proteins bind and transport cholesterol.Cell Metab. 2009 Jul;10(1):3-4. doi: 10.1016/j.cmet.2009.06.007. Cell Metab. 2009. PMID: 19583946
References
-
- Babalola JO, Wendeler M, Breiden B, Arenz C, Schwarzmann G, Locatelli-Hoops S, Sandhoff K. Development of an assay for the intermembrane transfer of cholesterol by Niemann-Pick C2 protein. Biol. Chem. 2007;388:617–626. - PubMed
-
- Brown MS, Goldstein JL. A receptor-mediated pathway for cholesterol homeostasis. Science. 1986;232:34–47. - PubMed
-
- Brown MS, Goldstein JL. The SREBP pathway: Regulation of cholesterol metabolism by proteolysis of a membrane-bound transcription factor. Cell. 1997;89:331–340. - PubMed
-
- Brunger AT, Adams PD, Clore GM, DeLano WL, Gros P, Grosse-Kunstleve RW, Jiang JS, Kuszewski J, Nilges M, Pannu NS, Read RJ, Rice LM, Simonson T, Warren GL. Crystallography & NMR system: A new software suite for macromolecular structure determination. Acta Crystallogr. D Biol. Crystallogr. 1998;D54:905–921. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Research Materials
