

The beneficial endophyte Trichoderma hamatum isolate DIS 219b promotes growth and delays the onset of the drought response in Theobroma cacao
- PMID: 19564160
- PMCID: PMC2718224
- DOI: 10.1093/jxb/erp165
The beneficial endophyte Trichoderma hamatum isolate DIS 219b promotes growth and delays the onset of the drought response in Theobroma cacao
Abstract
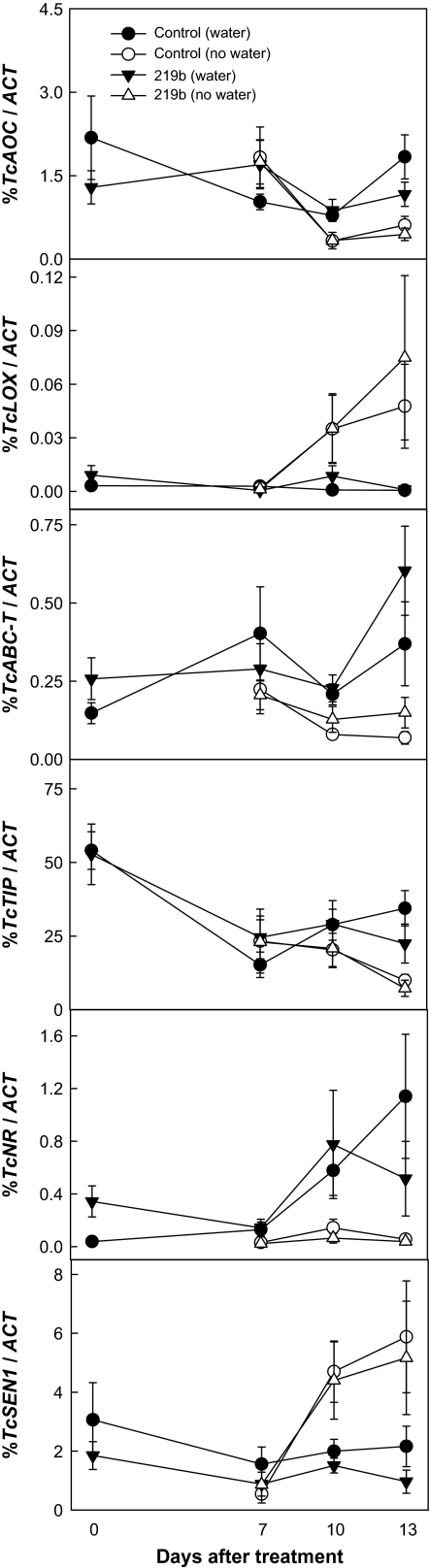
Theobroma cacao (cacao) is cultivated in tropical climates and is exposed to drought stress. The impact of the endophytic fungus Trichoderma hamatum isolate DIS 219b on cacao's response to drought was studied. Colonization by DIS 219b delayed drought-induced changes in stomatal conductance, net photosynthesis, and green fluorescence emissions. The altered expression of 19 expressed sequence tags (ESTs) (seven in leaves and 17 in roots with some overlap) by drought was detected using quantitative real-time reverse transcription PCR. Roots tended to respond earlier to drought than leaves, with the drought-induced changes in expression of seven ESTs being observed after 7 d of withholding water. Changes in gene expression in leaves were not observed until after 10 d of withholding water. DIS 219b colonization delayed the drought-altered expression of all seven ESTs responsive to drought in leaves by > or = 3 d, but had less influence on the expression pattern of the drought-responsive ESTs in roots. DIS 219b colonization had minimal direct influence on the expression of drought-responsive ESTs in 32-d-old seedlings. By contrast, DIS 219b colonization of 9-d-old seedlings altered expression of drought-responsive ESTs, sometimes in patterns opposite of that observed in response to drought. Drought induced an increase in the concentration of many amino acids in cacao leaves, while DIS 219b colonization caused a decrease in aspartic acid and glutamic acid concentrations and an increase in alanine and gamma-aminobutyric acid concentrations. With or without exposure to drought conditions, colonization by DIS 219b promoted seedling growth, the most consistent effects being an increase in root fresh weight, root dry weight, and root water content. Colonized seedlings were slower to wilt in response to drought as measured by a decrease in the leaf angle drop. The primary direct effect of DIS 219b colonization was promotion of root growth, regardless of water status, and an increase in water content which it is proposed caused a delay in many aspects of the drought response of cacao.
Figures







References
-
- Adams P, De-Leij FAAM, Lynch JM. Trichoderma harzianum Rifai 1295-22 mediates growth promotion of crack willow (Salix fragilis) saplings in both clean and metal-contaminated soil. Microbial Ecology. 2007;54:306–313. - PubMed
-
- Augé RM. Water relations, drought and VA mycorrhizal symbiosis. Mycorrhiza. 2001;11:3–42.
-
- Bae H, Kim S-H, Kim MS, Sicher RC, Strem MD, Natarajan S, Bailey BA. The drought response of Theobroma cacao (cacao) and the regulation of genes involved in polyamine biosynthesis by drought and other stresses. Plant Physiology and Biochemistry. 2008;46:174–188. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
