

Regulation of TLR7/9 responses in plasmacytoid dendritic cells by BST2 and ILT7 receptor interaction
- PMID: 19564354
- PMCID: PMC2715090
- DOI: 10.1084/jem.20090547
Regulation of TLR7/9 responses in plasmacytoid dendritic cells by BST2 and ILT7 receptor interaction
Abstract
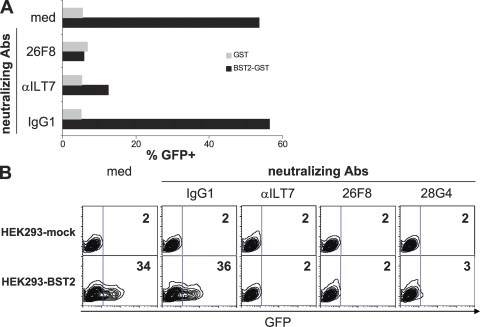
Plasmacytoid dendritic cells (pDCs) produce copious type I interferon (IFN) upon sensing nucleic acids through Toll-like receptor (TLR) 7 and TLR9. Uncontrolled pDC activation and IFN production are implicated in lymphopenia and autoimmune diseases; therefore, a mechanism controlling pDC IFN production is essential. Human pDCs specifically express an orphan receptor, immunoglobulin-like transcript 7 (ILT7). Here, we discovered an ILT7 ligand expressed by human cell lines and identified it as bone marrow stromal cell antigen 2 (BST2; CD317). BST2 directly binds to purified ILT7 protein, initiates signaling via the ILT7-FcepsilonRIgamma complex, and strongly inhibits production of IFN and proinflammatory cytokines by pDCs. Readily induced by IFN and other proinflammatory cytokines, BST2 may modulate the human pDC's IFN responses through ILT7 in a negative feedback fashion.
Figures






Similar articles
-
Signaling and ligand interaction of ILT7: receptor-mediated regulatory mechanisms for plasmacytoid dendritic cells.Immunol Rev. 2010 Mar;234(1):163-76. doi: 10.1111/j.0105-2896.2009.00867.x. Immunol Rev. 2010. PMID: 20193018 Free PMC article. Review.
-
Ig-like transcript 7, but not bone marrow stromal cell antigen 2 (also known as HM1.24, tetherin, or CD317), modulates plasmacytoid dendritic cell function in primary human blood leukocytes.J Immunol. 2013 Mar 15;190(6):2622-30. doi: 10.4049/jimmunol.1202391. Epub 2013 Feb 11. J Immunol. 2013. PMID: 23401591 Free PMC article.
-
Vpu Exploits the Cross-Talk between BST2 and the ILT7 Receptor to Suppress Anti-HIV-1 Responses by Plasmacytoid Dendritic Cells.PLoS Pathog. 2015 Jul 14;11(7):e1005024. doi: 10.1371/journal.ppat.1005024. eCollection 2015 Jul. PLoS Pathog. 2015. PMID: 26172439 Free PMC article.
-
Activation of the ILT7 receptor and plasmacytoid dendritic cell responses are governed by structurally-distinct BST2 determinants.J Biol Chem. 2019 Jul 5;294(27):10503-10518. doi: 10.1074/jbc.RA119.008481. Epub 2019 May 22. J Biol Chem. 2019. PMID: 31118237 Free PMC article.
-
Unraveling the functions of plasmacytoid dendritic cells during viral infections, autoimmunity, and tolerance.Immunol Rev. 2010 Mar;234(1):142-62. doi: 10.1111/j.0105-2896.2009.00881.x. Immunol Rev. 2010. PMID: 20193017 Free PMC article. Review.
Cited by
-
Vpu and BST2: Still Not There Yet?Front Microbiol. 2012 Apr 9;3:131. doi: 10.3389/fmicb.2012.00131. eCollection 2012. Front Microbiol. 2012. PMID: 22509177 Free PMC article.
-
The multifaceted biology of plasmacytoid dendritic cells.Nat Rev Immunol. 2015 Aug;15(8):471-85. doi: 10.1038/nri3865. Epub 2015 Jul 10. Nat Rev Immunol. 2015. PMID: 26160613 Free PMC article. Review.
-
Bone marrow stromal cell antigen 2: Tumor biology, signaling pathway and therapeutic targeting (Review).Oncol Rep. 2024 Mar;51(3):45. doi: 10.3892/or.2024.8704. Epub 2024 Jan 19. Oncol Rep. 2024. PMID: 38240088 Free PMC article. Review.
-
Cancer immunotherapy via dendritic cells.Nat Rev Cancer. 2012 Mar 22;12(4):265-77. doi: 10.1038/nrc3258. Nat Rev Cancer. 2012. PMID: 22437871 Free PMC article. Review.
-
Expansion of immunoglobulin-secreting cells and defects in B cell tolerance in Rag-dependent immunodeficiency.J Exp Med. 2010 Jul 5;207(7):1541-54. doi: 10.1084/jem.20091927. Epub 2010 Jun 14. J Exp Med. 2010. PMID: 20547827 Free PMC article.
References
-
- Arase H., Mocarski E.S., Campbell A.E., Hill A.B., Lanier L.L. 2002. Direct recognition of cytomegalovirus by activating and inhibitory NK cell receptors.Science. 296:1323–1326 - PubMed
-
- Bauer S., Groh V., Wu J., Steinle A., Phillips J.H., Lanier L.L., Spies T. 1999. Activation of NK cells and T cells by NKG2D, a receptor for stress-inducible MICA.Science. 285:727–729 - PubMed
-
- Blanco P., Palucka A.K., Gill M., Pascual V., Banchereau J. 2001. Induction of dendritic cell differentiation by IFN-alpha in systemic lupus erythematosus.Science. 294:1540–1543 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
