

Direct and indirect roles of RECQL4 in modulating base excision repair capacity
- PMID: 19567405
- PMCID: PMC2729667
- DOI: 10.1093/hmg/ddp291
Direct and indirect roles of RECQL4 in modulating base excision repair capacity
Abstract
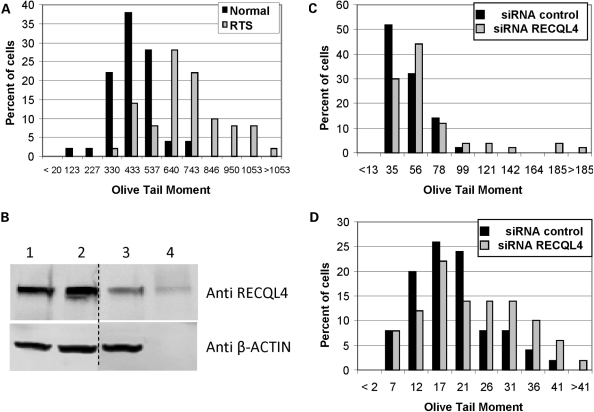
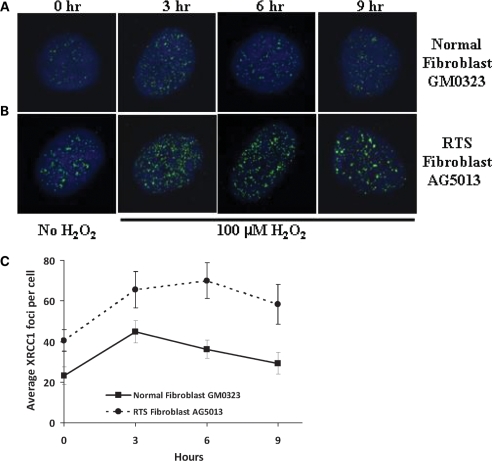
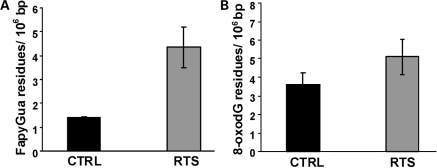
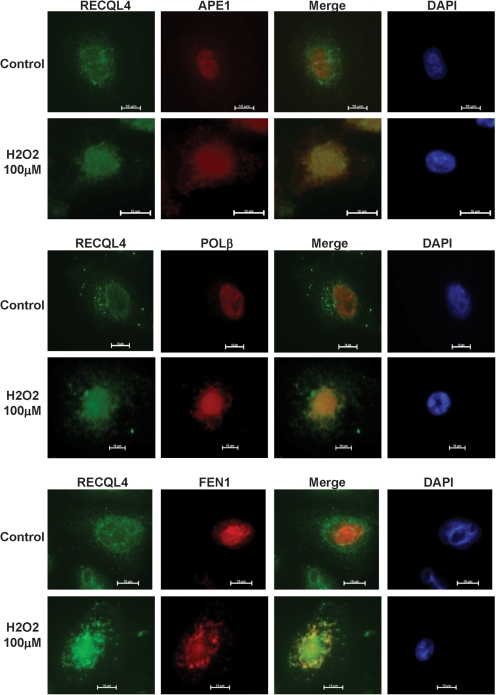
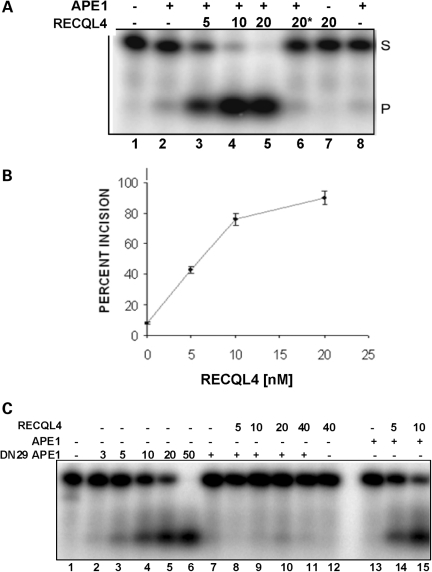
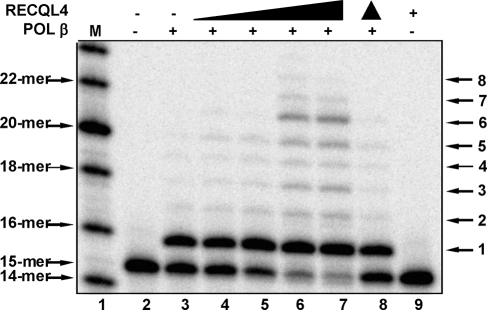
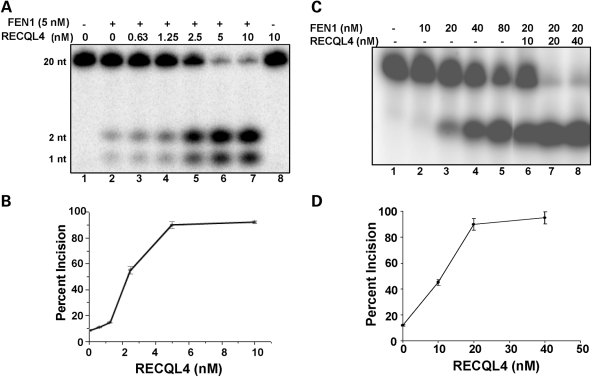
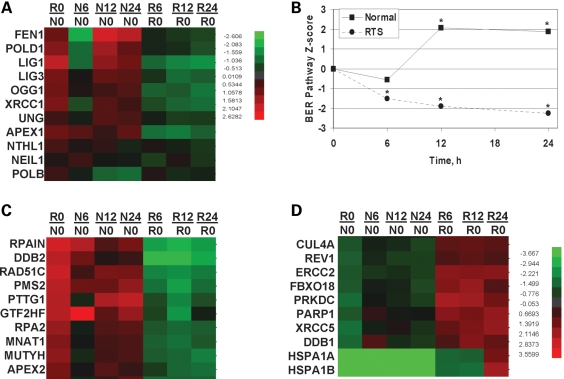
RECQL4 is a human RecQ helicase which is mutated in approximately two-thirds of individuals with Rothmund-Thomson syndrome (RTS), a disease characterized at the cellular level by chromosomal instability. BLM and WRN are also human RecQ helicases, which are mutated in Bloom and Werner's syndrome, respectively, and associated with chromosomal instability as well as premature aging. Here we show that primary RTS and RECQL4 siRNA knockdown human fibroblasts accumulate more H(2)O(2)-induced DNA strand breaks than control cells, suggesting that RECQL4 may stimulate repair of H(2)O(2)-induced DNA damage. RTS primary fibroblasts also accumulate more XRCC1 foci than control cells in response to endogenous or induced oxidative stress and have a high basal level of endogenous formamidopyrimidines. In cells treated with H(2)O(2), RECQL4 co-localizes with APE1, and FEN1, key participants in base excision repair. Biochemical experiments indicate that RECQL4 specifically stimulates the apurinic endonuclease activity of APE1, the DNA strand displacement activity of DNA polymerase beta, and incision of a 1- or 10-nucleotide flap DNA substrate by Flap Endonuclease I. Additionally, RTS cells display an upregulation of BER pathway genes and fail to respond like normal cells to oxidative stress. The data herein support a model in which RECQL4 regulates both directly and indirectly base excision repair capacity.
Figures
References
-
- Wang L.L., Levy M.L., Lewis R.A., Chintagumpala M.M., Lev D., Rogers M., Plon S.E. Clinical manifestations in a cohort of 41 Rothmund-Thomson syndrome patients. Am. J. Med. Genet. 2001;102:11–17. - PubMed
-
- Wang L.L., Gannavarapu A., Kozinetz C.A., Levy M.L., Lewis R.A., Chintagumpala M.M., Ruiz-Maldanado R., Contreras-Ruiz J., Cunniff C., Erickson R.P., et al. Association between osteosarcoma and deleterious mutations in the RECQL4 gene in Rothmund-Thomson syndrome. J. Natl Cancer Inst. 2003;95:669–674. - PubMed
-
- Siitonen H.A., Kopra O., Kaariainen H., Haravuori H., Winter R.M., Saamanen A.M., Peltonen L., Kestila M. Molecular defect of RAPADILINO syndrome expands the phenotype spectrum of RECQL diseases. Hum. Mol. Genet. 2003;12:2837–2844. - PubMed
-
- Van Maldergem L., Siitonen H.A., Jalkh N., Chouery E., De Roy M., Delague V., Muenke M., Jabs E.W., Cai J., Wang L.L., et al. Revisiting the craniosynostosis-radial ray hypoplasia association: Baller-Gerold syndrome caused by mutations in the RECQL4 gene. J. Med. Genet. 2006;43:148–152. - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
