

Hsp90 is an essential regulator of EphA2 receptor stability and signaling: implications for cancer cell migration and metastasis
- PMID: 19567782
- PMCID: PMC3155284
- DOI: 10.1158/1541-7786.MCR-08-0582
Hsp90 is an essential regulator of EphA2 receptor stability and signaling: implications for cancer cell migration and metastasis
Abstract
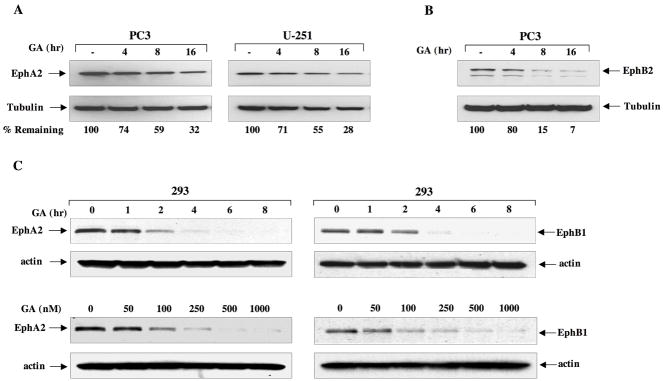
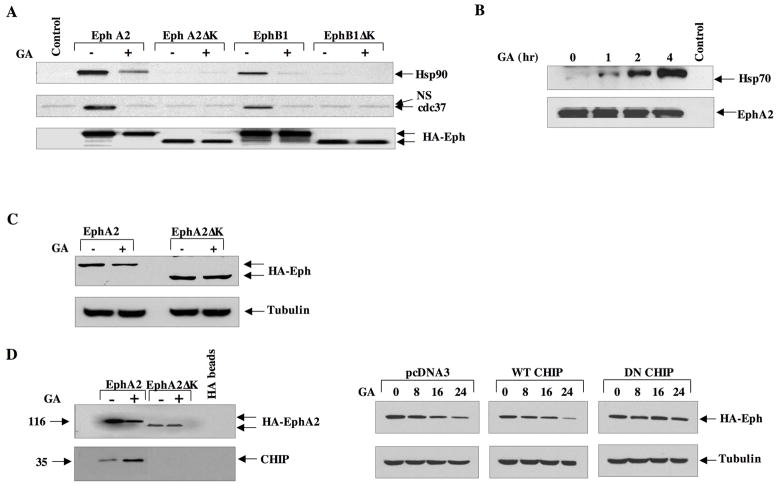
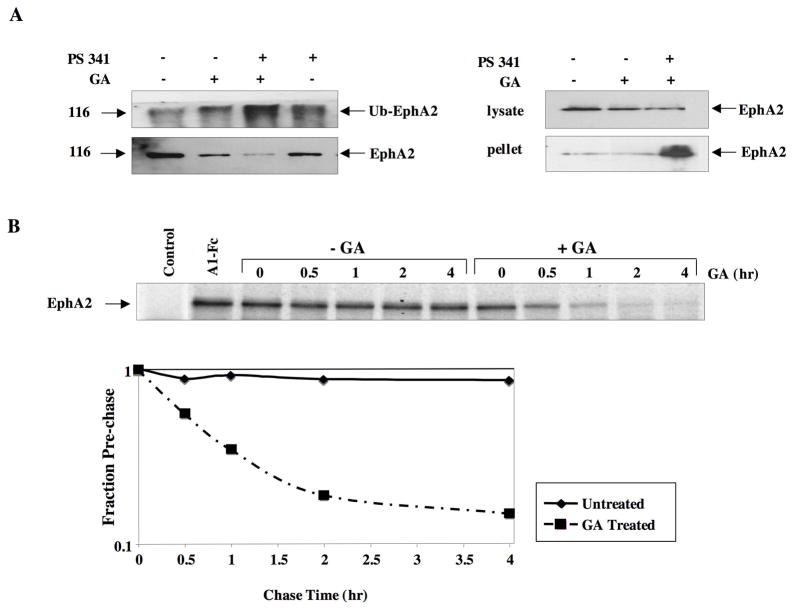
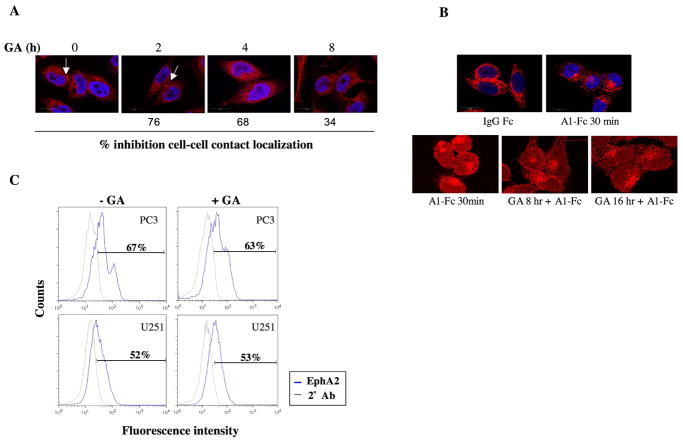
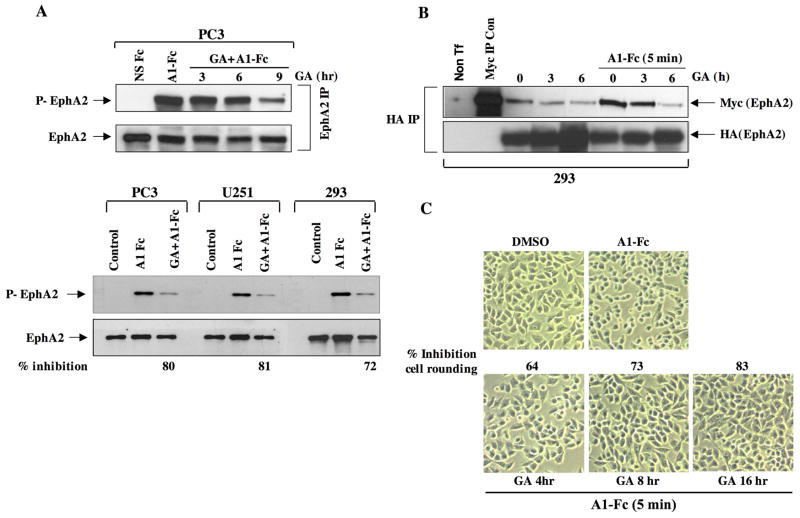
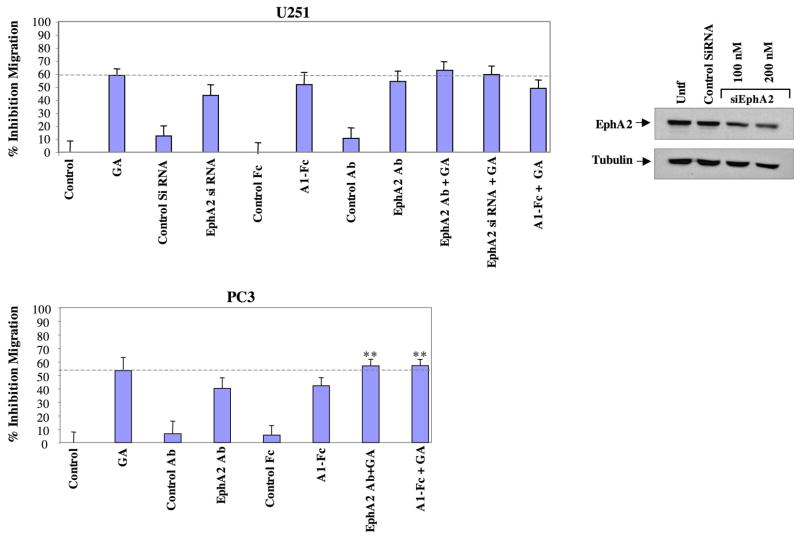
A subset of Eph receptors and their corresponding ligands are commonly expressed in tumor cells where they mediate biological processes such as cell migration and adhesion, whereas their expression in endothelial cells promotes angiogenesis. In particular, the tumor-specific up-regulation of EphA2 confers properties of increased cellular motility, invasiveness, tumor angiogenesis, and tumor progression, and its overexpression correlates with poor prognosis in several cancer types. The cellular chaperone Hsp90 also plays a significant role in regulating cell migration and angiogenesis, although the full repertoire of motility driving proteins dependent on Hsp90 function remain poorly defined. We explored the hypothesis that Hsp90 may regulate the activity of EphA2 and examined the potential relationship between EphA2 receptor signaling and chaperone function. We show that geldanamycin, an Hsp90 antagonist, dramatically destabilizes newly synthesized EphA2 protein and diminishes receptor levels in a proteasome-dependent pathway. In addition, geldanamycin treatment impairs EphA2 signaling, as evidenced by a decrease in ligand-dependent receptor phosphorylation and subsequent cell rounding. Therefore, Hsp90 exerts a dual role in regulating the stability of nascent EphA2 protein and maintaining the signaling capacity of the mature receptor. Our findings also suggest that the geldanamycin-dependent mitigation of EphA2 signaling in receptor-overexpressing cancer cells may be sufficient to recapitulate the antimotility effects of this drug. Finally, the identification of a pharmacologic approach to suppress EphA2 expression and signaling highlights the attractive possibility that Hsp90 inhibitors may have clinical utility in antagonizing EphA2-dependent tumorigenic progression.
Figures
References
-
- Lauffenburger DA, Horwitz AF. Cell migration: a physically integrated molecular process. Cell. 1996;84:359–369. - PubMed
-
- Pasquale EB. Eph-ephrin bidirectional signaling in physiology and disease. Cell. 2008;133:38–52. - PubMed
-
- Pasquale EB. Eph receptor signalling casts a wide net on cell behaviour. Nat Rev Mol Cell Biol. 2005;6:462–475. - PubMed
-
- Gale NW, Holland SJ, Valenzuela DM, Flenniken A, Pan L, Ryan TE, et al. Eph receptors and ligands comprise two major specificity subclasses and are reciprocally compartmentalized during embryogenesis. Neuron. 1996;17:9–19. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
