

Amino acid position-specific contributions to amyloid beta-protein oligomerization
- PMID: 19567875
- PMCID: PMC2749133
- DOI: 10.1074/jbc.M109.038133
Amino acid position-specific contributions to amyloid beta-protein oligomerization
Abstract
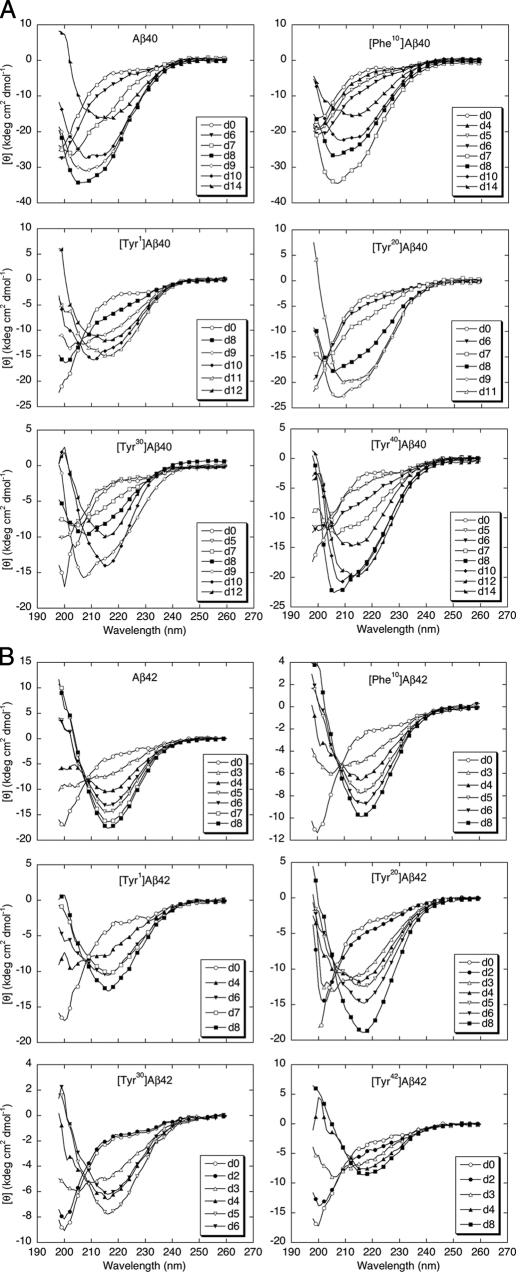
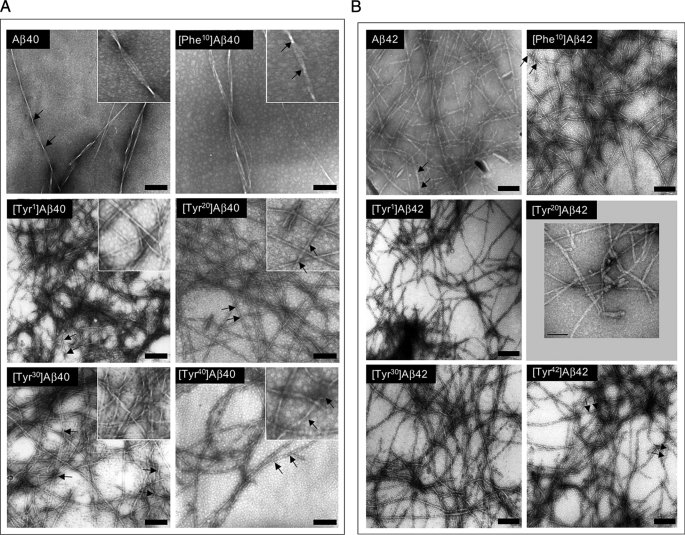
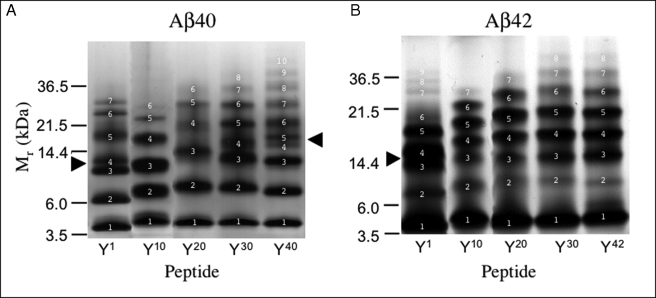
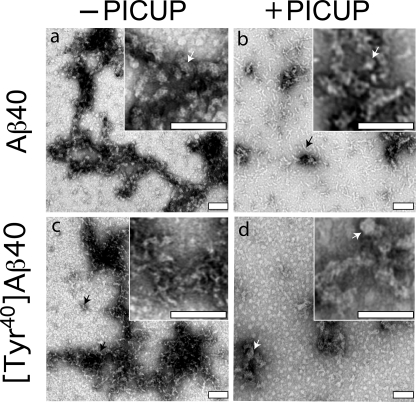
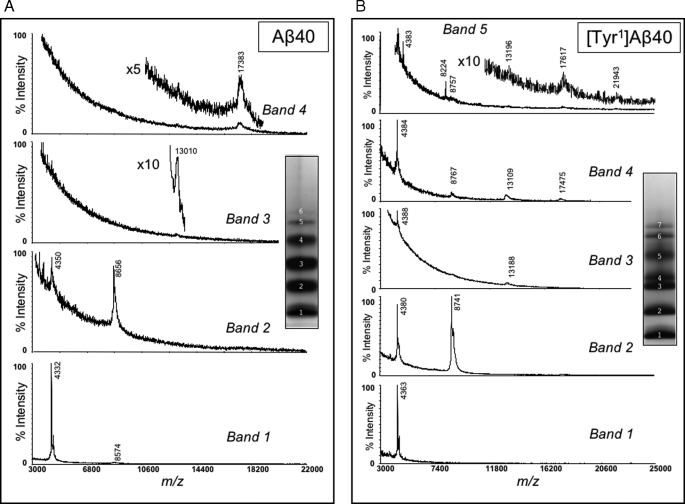
Understanding the structural and assembly dynamics of the amyloid beta-protein (Abeta) has direct relevance to the development of therapeutic agents for Alzheimer disease. To elucidate these dynamics, we combined scanning amino acid substitution with a method for quantitative determination of the Abeta oligomer frequency distribution, photo-induced cross-linking of unmodified proteins (PICUP), to perform "scanning PICUP." Tyr, a reactive group in PICUP, was substituted at position 1, 10, 20, 30, or 40 (for Abeta40) or 42 (for Abeta42). The effects of these substitutions were probed using circular dichroism spectroscopy, thioflavin T binding, electron microscopy, PICUP, and mass spectrometry. All peptides displayed a random coil --> alpha/beta --> beta transition, but substitution-dependent alterations in assembly kinetics and conformer complexity were observed. Tyr(1)-substituted homologues of Abeta40 and Abeta42 assembled the slowest and yielded unusual patterns of oligomer bands in gel electrophoresis experiments, suggesting oligomer compaction had occurred. Consistent with this suggestion was the observation of relatively narrow [Tyr(1)]Abeta40 fibrils. Substitution of Abeta40 at the C terminus decreased the population conformational complexity and substantially extended the highest order of oligomers observed. This latter effect was observed in both Abeta40 and Abeta42 as the Tyr substitution position number increased. The ability of a single substitution (Tyr(1)) to alter Abeta assembly kinetics and the oligomer frequency distribution suggests that the N terminus is not a benign peptide segment, but rather that Abeta conformational dynamics and assembly are affected significantly by the competition between the N and C termini to form a stable complex with the central hydrophobic cluster.
Figures

References
-
- Hebert L. E., Scherr P. A., Bienias J. L., Bennett D. A., Evans D. A. (2003) Arch. Neurol. 60,1119–1122 - PubMed
-
- Brookmeyer R., Johnson E., Ziegler-Graham K., Arrighi H. M. (2007) Alzheimers Dement. 3,186–191 - PubMed
-
- Kirkitadze M. D., Bitan G., Teplow D. B. (2002) J. Neurosci. Res. 69,567–577 - PubMed
-
- Klein W. L., Stine W. B., Jr., Teplow D. B. (2004) Neurobiol. Aging 25,569–580 - PubMed
-
- Klyubin I., Walsh D. M., Cullen W. K., Fadeeva J. V., Anwyl R., Selkoe D. J., Rowan M. J. (2004) Eur. J. Neurosci. 19,2839–2846 - PubMed
