

Development of kidney glomerular endothelial cells and their role in basement membrane assembly
- PMID: 19568349
- PMCID: PMC2659369
- DOI: 10.4161/org.7577
Development of kidney glomerular endothelial cells and their role in basement membrane assembly
Abstract
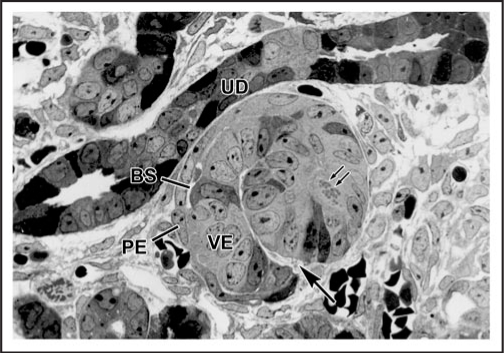
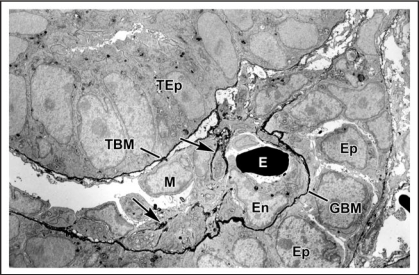
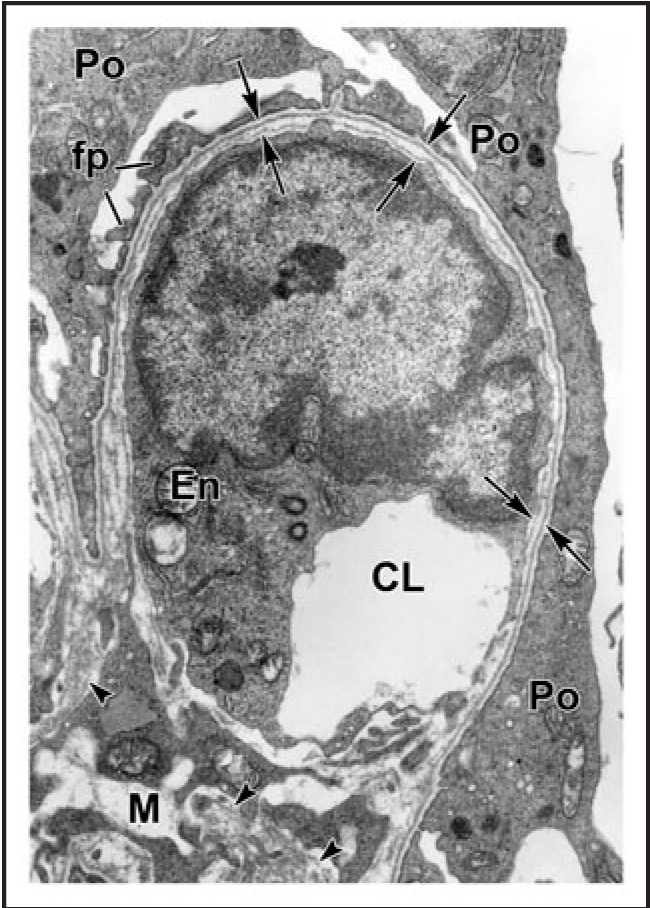
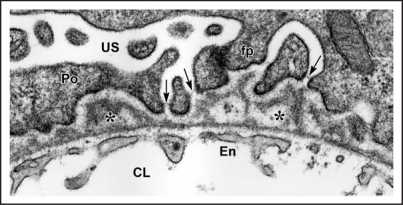
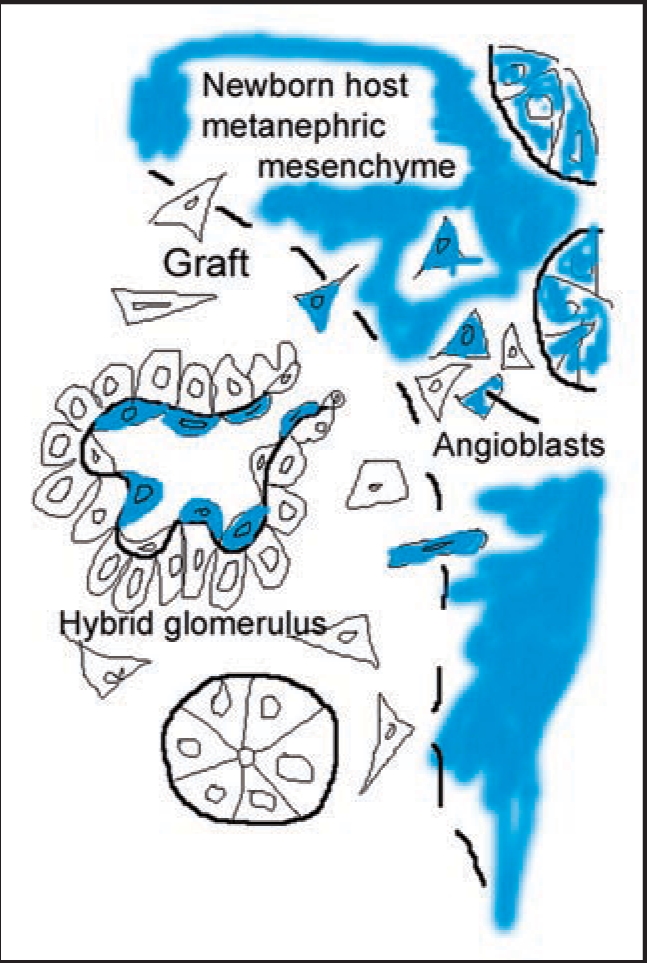
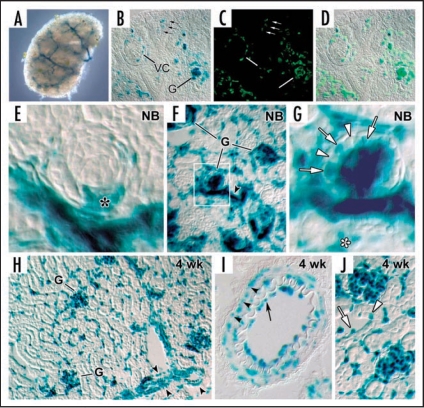
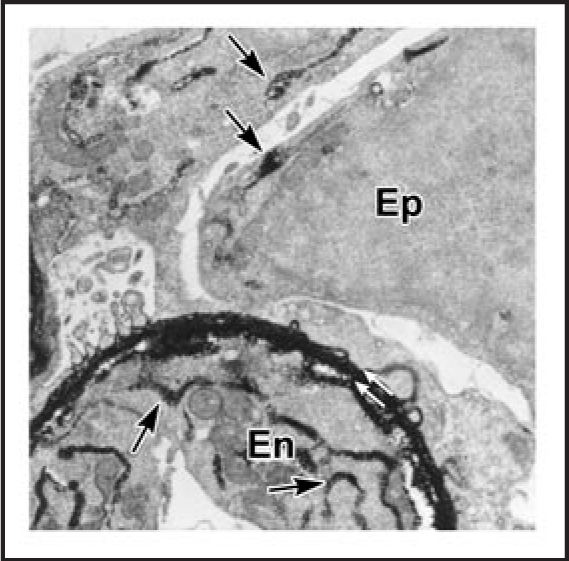
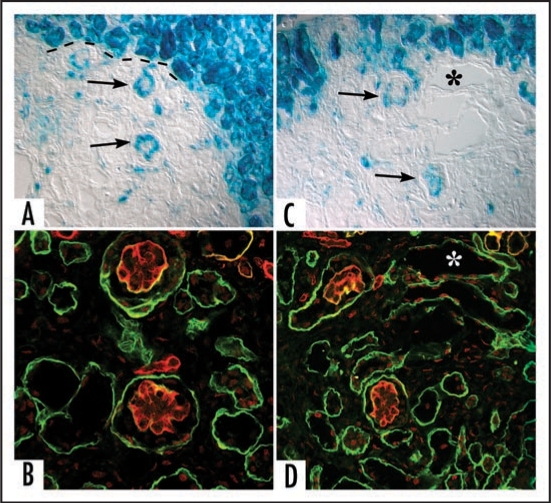
Data showing that the embryonic day 12 (E12) mouse kidney contains its own pool of endothelial progenitor cells is presented. Mechanisms that regulate metanephric endothelial recruitment and differentiation, including the hypoxia-inducible transcription factors and vascular endothelial growth factor/vascular endothelial growth factor receptor signaling system, are also discussed. Finally, evidence that glomerular endothelial cells contribute importantly to assembly of the glomerular basement membrane (GBM), especially the laminin component, is reviewed. Together, this forum offers insights on blood vessel development in general, and formation of the glomerular capillary in particular, which inarguably is among the most unique vascular structures in the body.
Keywords: glomerular basement membrane; laminin; podocytes; type IV collagen; vascular endothelial growth factor.
Figures
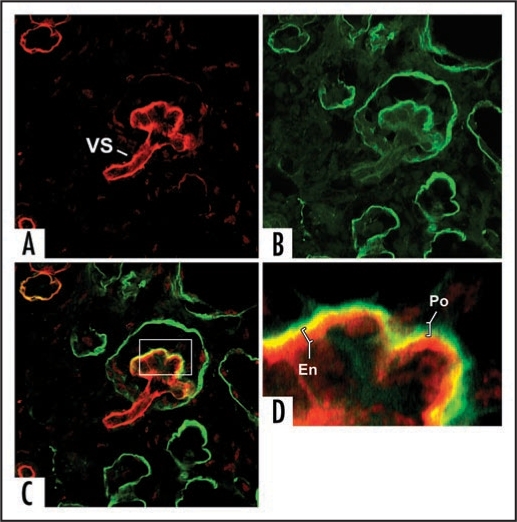
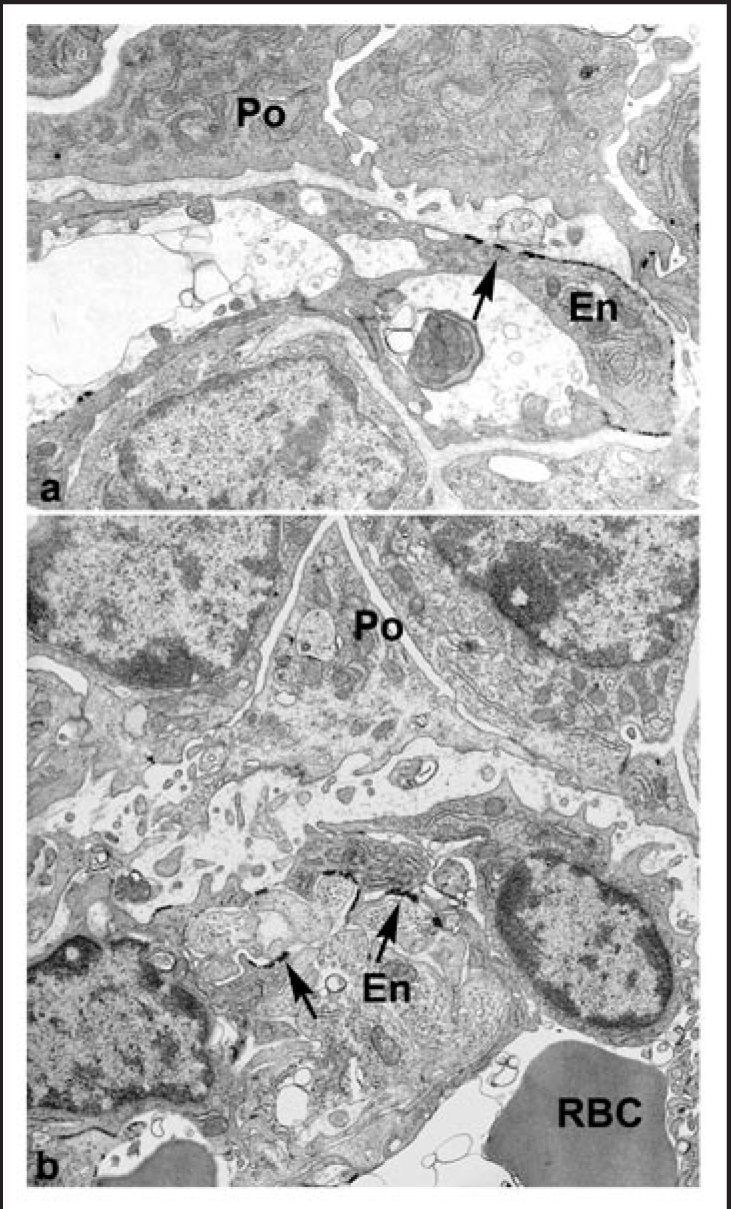
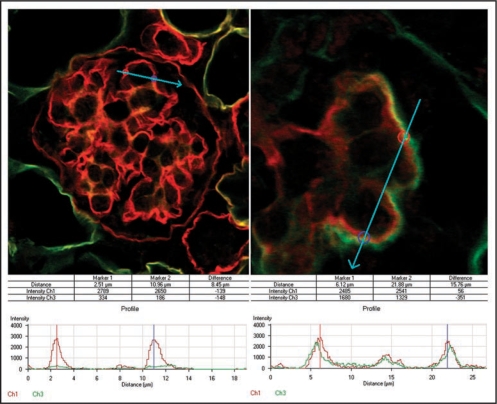
References
-
- Jeansson M, Haraldsson B. Morphological and functional evidence for an important role of the endothelial cell glycocalyx in the glomerular barrier. Am J Physiol Renal Physiol. 2006;290:111–116. - PubMed
-
- Somlo S, Mundel P. Getting a foothold in nephrotic syndrome. Nat Genet. 2000;24:333–335. - PubMed
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources