

Intracellular localization and conformational state of transglutaminase 2: implications for cell death
- PMID: 19568436
- PMCID: PMC2701606
- DOI: 10.1371/journal.pone.0006123
Intracellular localization and conformational state of transglutaminase 2: implications for cell death
Abstract
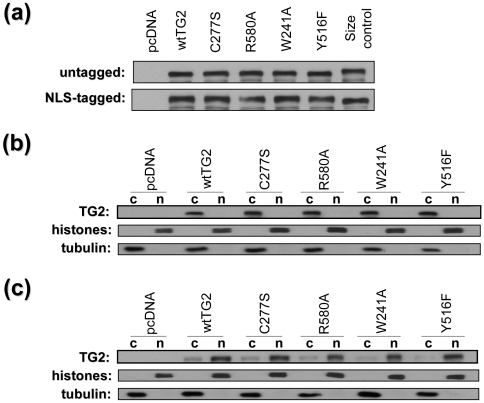
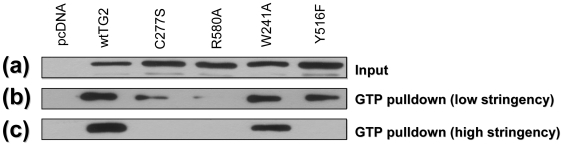
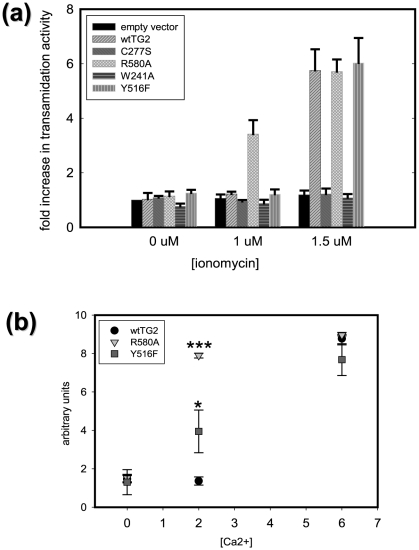
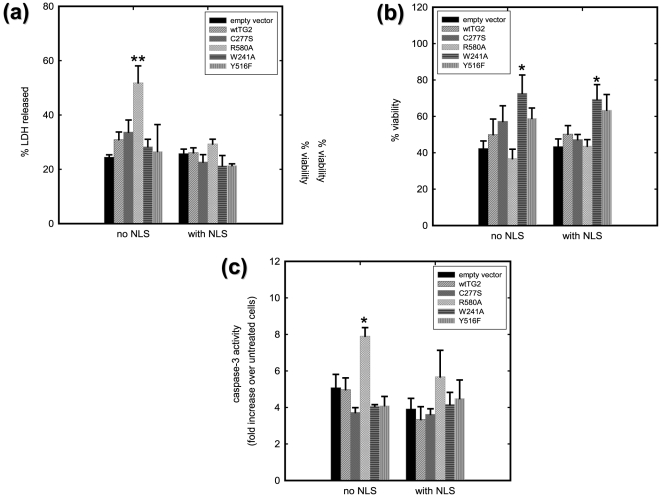
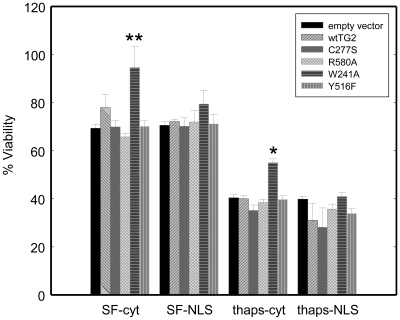
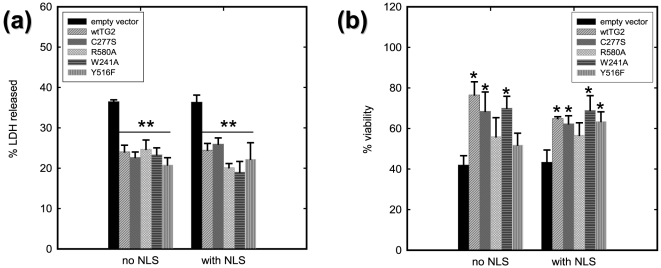
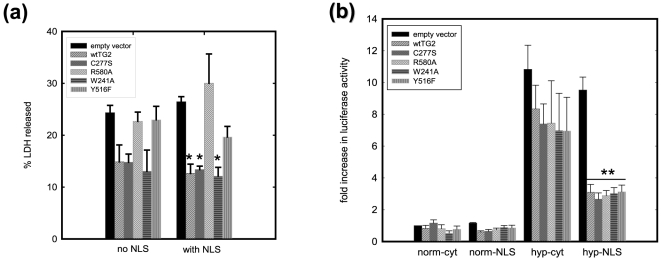
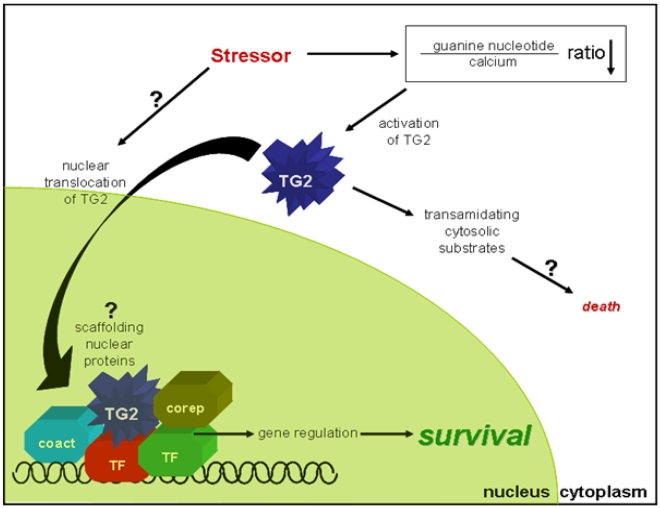
Transglutaminase 2 (TG2) is a multifunctional enzyme that has guanine nucleotide binding and GTP hydrolyzing activity in addition to its transamidating function. Studies show that TG2 is a player in mediating cell death processes. However, there is far from a consensus about the role of this enzyme in cell death processes as it appears to be dependent upon the cell type, stimuli, subcellular localization and conformational state of the enzyme. The purpose of this study was to dissect the role of TG2 in the cell death processes. To this end, we created and characterized 4 distinct point mutants of TG2, each of which differs from the wild type by its conformation or by lacking an important function. We also prepared these mutants as nuclear targeted proteins. By overexpressing mutant or wild type forms of TG2 in HEK 293 cells, we investigated the modulatory role of the protein in the cell death process in response to three stressors: thapsigargin, hyperosmotic stress and oxygen/glucose deprivation (OGD). All of the TG2 constructs, except the R580A mutant (which cannot bind guanine nucleotides and is therefore more prone to exhibit transamidating activity), either did not significantly affect the cell death processes or were protective. However in the case of the R580A mutant, cell death in response to high thapsigargin concentrations, was significantly increased. Intriguingly, nuclear localization of R580A-TG2 was sufficient to counteract the pro-death role of cytoplasmic R580A-TG2. In addition, nuclear localization of TG2 significantly facilitated its protective role against OGD. Our data support the hypothesis that the transamidation activity of TG2, which is mostly quiescent except in extreme stress conditions, is necessary for its pro-death role. In addition, nuclear localization of TG2 generally plays a key role in its protective function against cell death processes, either counteracting the detrimental effect or strengthening the protective role of the protein.
Conflict of interest statement
Figures
References
-
- Chen JS, Mehta K. Tissue transglutaminase: an enzyme with a split personality. Int J Biochem Cell Biol. 1999;31:817–836. - PubMed
-
- Nakaoka H, Perez DM, Baek KJ, Das T, Husain A, et al. Gh: a GTP-binding protein with transglutaminase activity and receptor signaling function. Science. 1994;264:1593–1596. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
