

doi: 10.1007/s12154-008-0010-6.
Epub 2008 Sep 23.
Sterols and membrane dynamics
Affiliations
- PMID: 19568799
- PMCID: PMC2698314
- DOI: 10.1007/s12154-008-0010-6
Item in Clipboard
Sterols and membrane dynamics
J Chem Biol.
2008 Nov.
Abstract
The effect of sterols from mammals, plants, fungi, and bacteria on model and natural membrane dynamics are reviewed, in the frame of ordering-disordering properties of membranes. It is shown that all sterols share a common property: the ability to regulate dynamics in order to maintain membranes in a microfluid state where it can convey important biological processes. Depending on the sterol class, this property is modulated by molecular modifications that have occurred during evolution. The role of sterols in rafts, antibiotic complexes, and in protecting membranes from the destructive action of amphipathic toxins is also discussed.
Figures
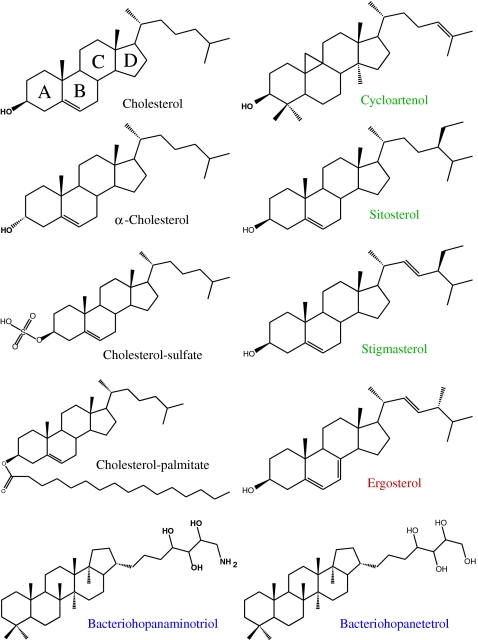
Molecular structures of mammalian, plant, fungus, and bacterial sterols
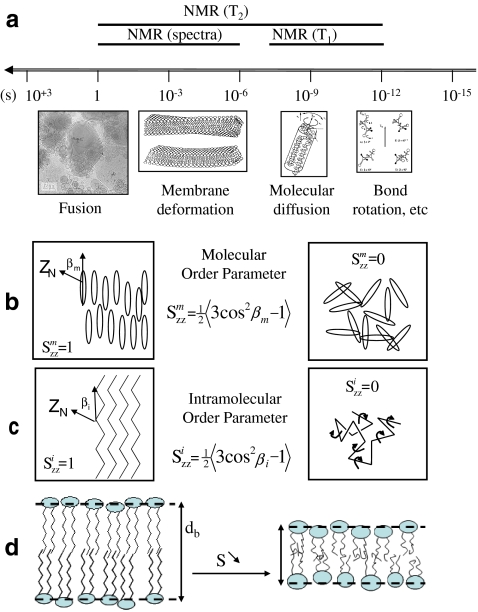
a Motional time scales in biomembranes and NMR windows. Drawings and microscopy image depict the spatial scale at which events may occur. Lower panel—order parameter concept. b Molecular order; c intramolecular (bonds) order; d correlation between order and membrane thickness, db—left, ordered membrane with little bond or molecule fluctuations (large db); right, less ordered membrane with bond and molecule fast reorientations within the membrane (small db). Adapted from [12]
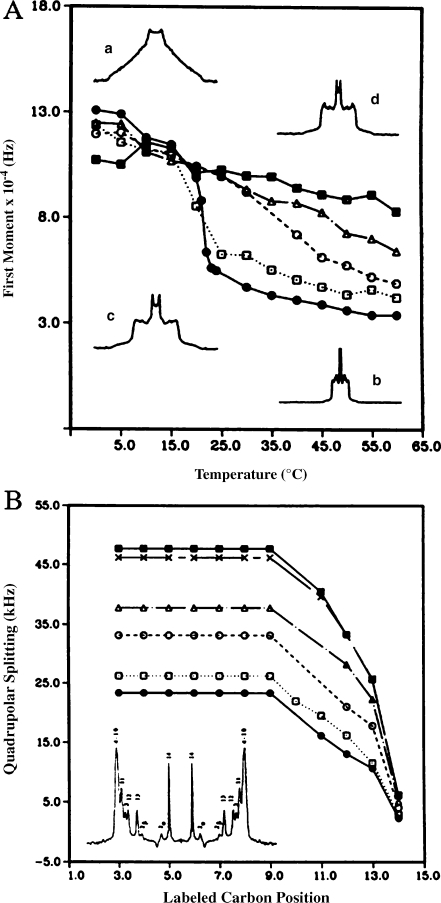
Temperature variation of the first moment (proportional to chain order parameter), M1, of 2H-labeled DMPC spectra in the presence of various amounts of cholesterol: pure lipid (filled circles), 10% (empty squares), 20% (empty circles), 30% (empty triangles), 50% (filled squares). Inserts show typical 2H-NMR powder spectra: pure lipid at 0 °C (a) and 60 °C (b), lipid plus 50% CHO at 0 °C (d) and 60 °C (c). b Quadrupolar splitting (proportional to bond order parameter) from de-Paked spectra (see insert) of [2H27-DMPC] as a function of labeled carbon position, at T = 50 °C. Same symbols as for a with in addition 40 mol% CHO (crosses). Numbers in the de-Paked spectrum (insert) represent the assignment of labeled carbon positions. Adapted from [42]

Spatial representation of the fused ring system of cholesterol and cholesteryl palmitate. The arrow indicates the direction of the rotation axis, n, collinear with the membrane normal. The symbol (empty circle) is the oxygen attached at C3 and deuterium nuclei of interest are plotted as small open circles. The palmitic chain in cholesteryl palmitate is attached at the oxygen. Molecules are drawn in the overall motional averaging axis system. The figure does not account for the ‘wobbling’ of the steroid skeleton (i.e., the drawing is made for Sm = 1). Adapted from [43]
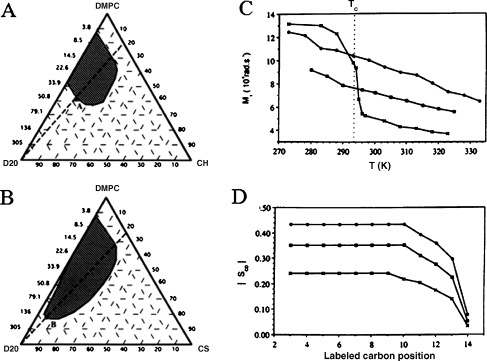
Partial phase diagrams of DMPC–CHO–D2O (a) and DMPC–CHS–D2O (b) at 25 °C. Hatched areas represent the lamellar phase domain. Compositions are given in weight percentages or corresponding D2O-to-DMPC molar ratio, Ri. Right panel—comparative evolution of the first spectral moment, M1 (chain order parameter), and the bond order parameter, SCD, for DMPC in the presence and absence of 30 mol% steroid. c Thermal evolution of M1, Tc is the DMPC main phase transition temperature. dSCD as a function of the labeled carbon position at 25 °C; DMPC (crosses), DMPC–CHO (30 mol%; filled circles), DMPC–CHS (30 mol%; filled squares). Adapted from [44]
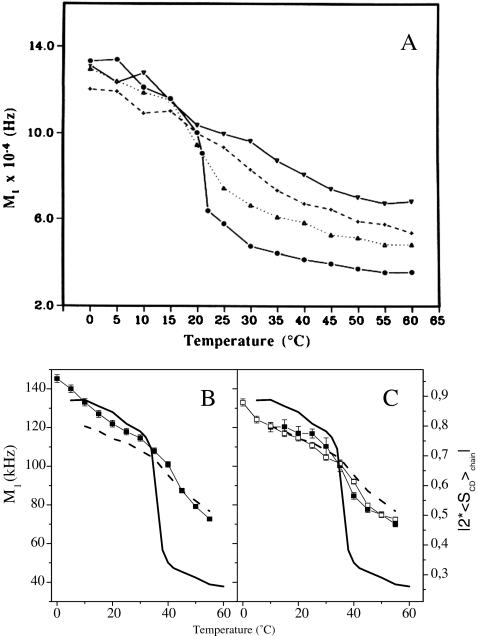
a Temperature variation of the first moment (proportional to chain order parameter), M1, of 2H-labeled DMPC spectra in the presence of various amounts of cycloartenol: pure lipid (filled circles), 10% (filled triangles), 20% (crosses), 30% (inverted filled triangles). Lower panel—first moments of 2H-NMR DPPC spectra with plant/fungi sterols versus temperature. First moments of pure DPPC-2H62 spectra (line without symbols) and of DPPC-2H62/CHO (30 mol%) spectra (dashed line without symbols) are shown in all diagrams for comparison. b DPPC-2H62 plus 30 mol% ERG (squares); c DPPC-2H62/STI (30 mol%; filled squares); DPPC-2H/SIT (30 mol%; empty squares). On double y-axis is plotted twice the chain order parameter. Adapted from [23] and [6]
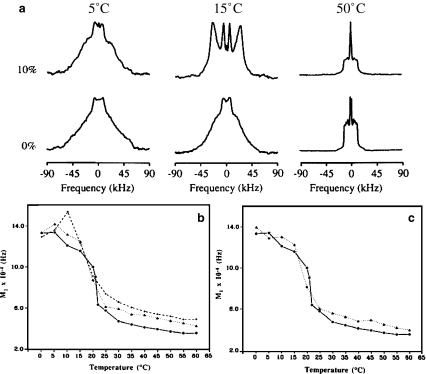
Hopanes and DMPC membranes. a2H27-DMPC spectra in the absence (lower row) and presence (upper row) of BHT (10 mol%). Lower panel—temperature variation of the first moment (proportional to chain order parameter), M1, of 2H-labeled DMPC spectra in the presence of various amounts of hopanes. Pure lipid (filled circles), b BHAT 10% (filled triangles), 20% (crosses), c BHT 10% (filled triangles)

aLeft—representation (stick mode) of cholesterol average orientation with respect to its principal axis of motional averaging, n (membrane normal), when n is parallel to the page plane. Right—view with n, perpendicular to the page plane, CPK representation is used here. The dashed line tentatively represents the cholesterol molecular area when viewed from above the membrane surface. b Thermal variation of the cholesterol molecular order parameter, Smol, in the four lipid–cholesterol compositions: POPC/CHO-2H5, SM/CHO-2H5, POPC/SM/CHO-2H5, SCRL/CHO-2H5. Adapted from [45]
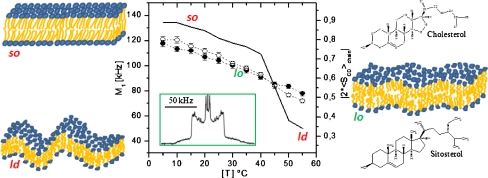
Regulation of temperature-driven membrane dynamics by plant sterols. Central panel—first spectral moment (left y-axis) or order parameter (right y-axis) as a function of temperature; solid line2H-DPPC with glucosylcerebroside; open circles plus stigmasterol; filled circles plus sitosterol. Insert2H-NMR spectrum typical of a liquid-ordered, lo, state. Left panel—schematics of solid-ordered, so (gel), and liquid-disordered, ld (fluid), membrane states. Right panel—schematics of the lo (raft) membrane state together with the structures of cholesterol and sitosterol. Adapted from [6, 7]
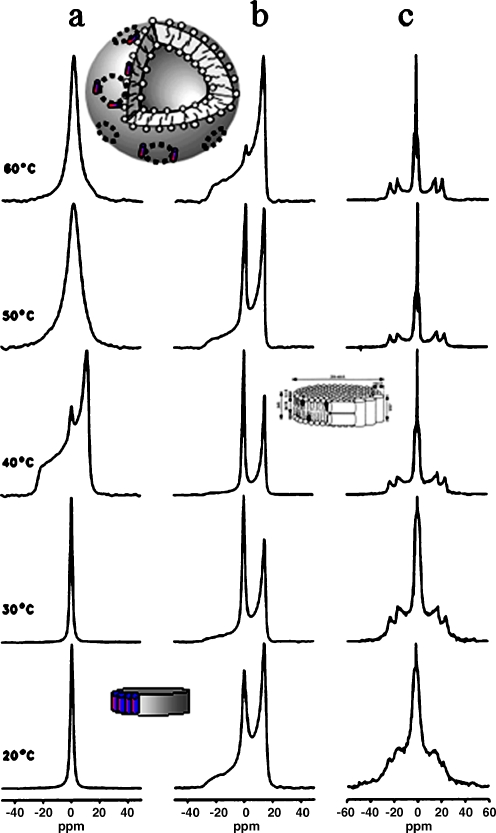
31P- and 2H-NMR spectra of the melittin/DPPC system in the presence and absence of cholesterol. a31P-NMR spectra for selected temperatures of the cholesterol-free system at lipid to melittin molar ratio of 20. b The corresponding 31P-NMR and c2H-NMR spectra of the system containing 30 mol% of 2H-labeled cholesterol. Inserts show the DPPC/melittin discs at low temperatures and the large vesicles at high temperatures. In the presence of CHO, remaining discs contain small amounts of sterol (insert on right hand side). Adapted from [36]

a Temperature dependence of the relaxation time, T1z, of [6-2H]-CHO in DMPC, in the presence or absence of amphotericin B (Ampho B). Lower panels—2H-NMR spectra of [2,2,3,4,4,6-2H6]cholesterol in DMPC (30 mol%) in the presence (b) and absence (c) of filipin, at 25 °C. The antibiotic, when present, is equimolar to cholesterol. Adapted from [38]
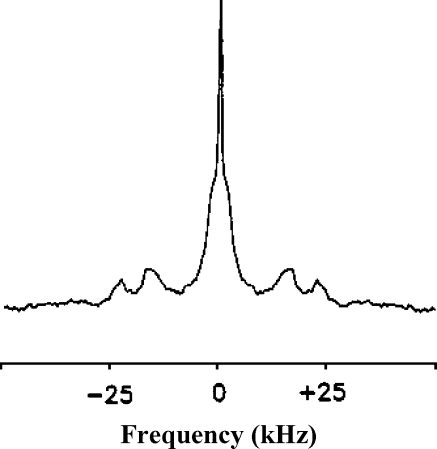
2H-NMR spectrum of [2,2,3,4,4,6-2H]cholesterol in membranes of human erythrocytes. Adapted from [39]
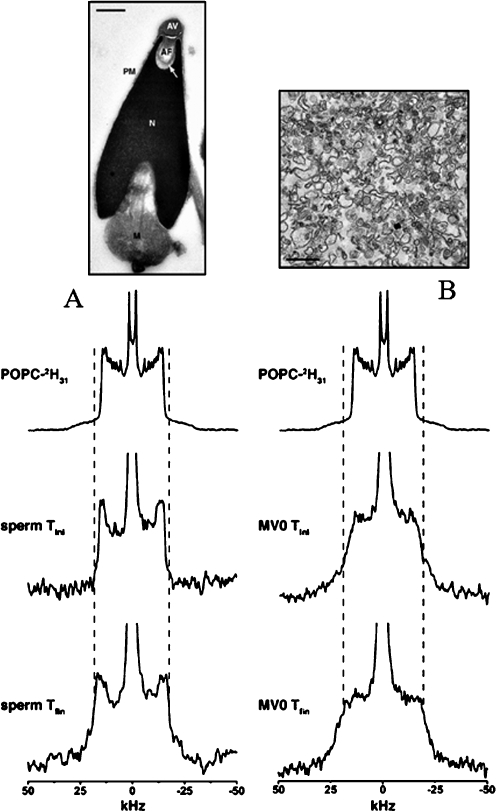
Deuterium solid-state NMR spectra of POPC-2H31 labeled sea urchin sperm membranes (a) and precursor egg membrane vesicles (MV0; b). Sperm and MV0 were labeled with MLVs and SUVs of POPC respectively for 30 min at 40 °C and the corresponding deuterium NMR spectra acquired at 10 °C (middle spectra «sperm Tini», «MV0 Tini»). The systems were equilibrated at 40 °C and reacquired at 10 °C (bottom spectra «sperm Tfin», «MV0 Tfin»). The top spectra corresponds to MLVs of POPC and the dashed lines show the plateau quadrupolar splitting enlargement on sperm and MV0 spectra postequilibrium at 40 °C. Upper images are electron microscopy pictures of sperm and egg membrane vesicles. Adapted from [40]
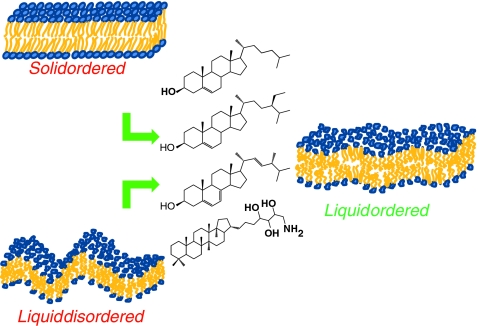
Sterols (mammals, fungi, plants, bacteria) as regulators of membrane dynamics. Left—the solid-ordered and liquid-disordered states in the absence of sterols. Right—the liquid ordered state with sterols
References
-
- Ribeiro N, Streiff S, Heissler D, Elhabiri M, Albrecht-Gary AM, Atsumi M, Gotoh M, Desaubry L, Nakatani Y, Ourisson G. Reinforcing effect of bi- and tri-cyclopolyprenols on ‘primitive’ membranes made of polyprenyl phosphates. Tetrahedron. 2007;63:3395–3407. doi: 10.1016/j.tet.2007.01.076. - DOI
-
- Saito H, Suzuki N. Distributions and sources of hopanes, hopanoic acids and hopanols in Miocene to recent sediments from ODP Leg 190, Nankai Trough. Org Geochem. 2007;38:1715–1728. doi: 10.1016/j.orggeochem.2007.05.012. - DOI
LinkOut - more resources
Full Text Sources
Other Literature Sources
