

TWIK-1 and TREK-1 are potassium channels contributing significantly to astrocyte passive conductance in rat hippocampal slices
- PMID: 19571146
- PMCID: PMC6665656
- DOI: 10.1523/JNEUROSCI.5784-08.2009
TWIK-1 and TREK-1 are potassium channels contributing significantly to astrocyte passive conductance in rat hippocampal slices
Abstract
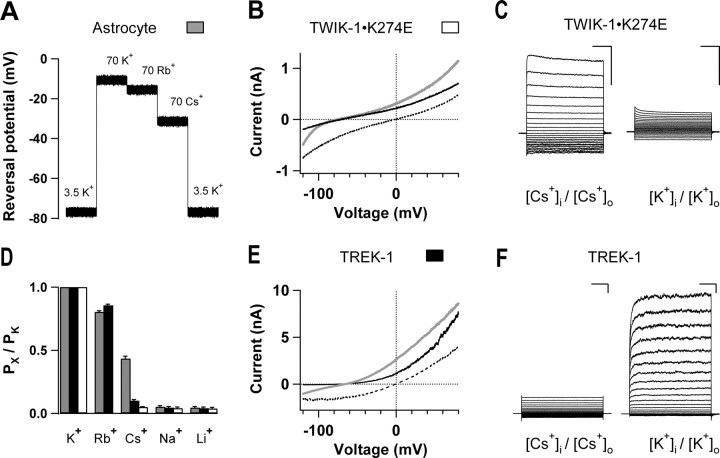
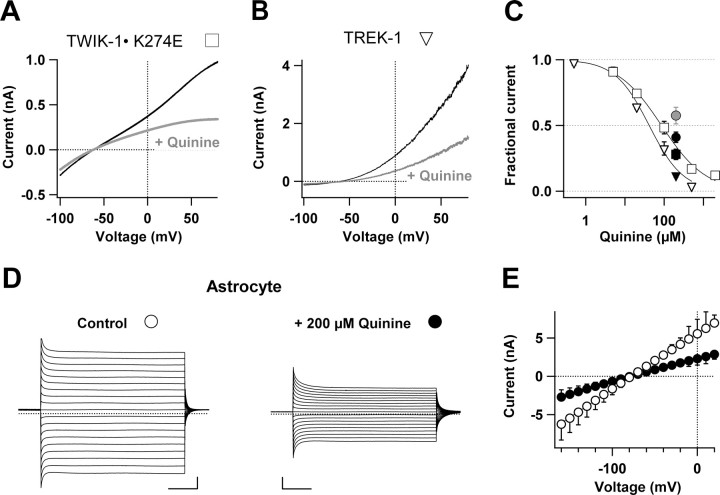
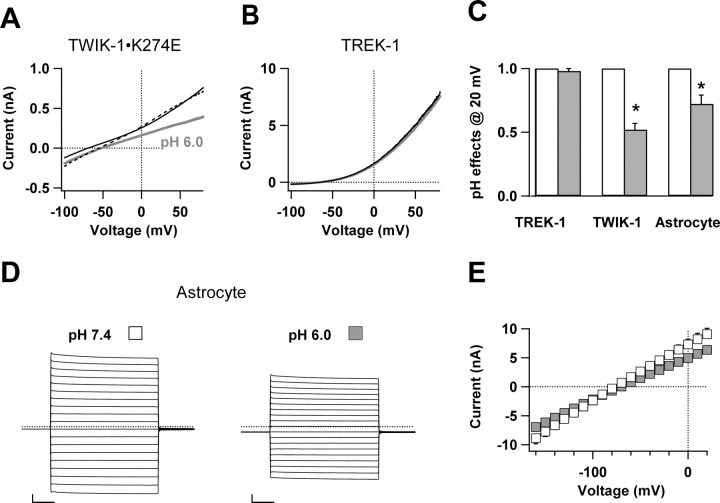
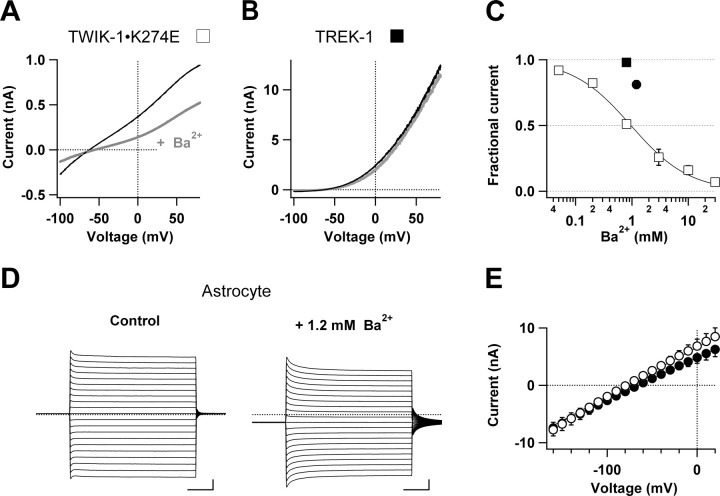
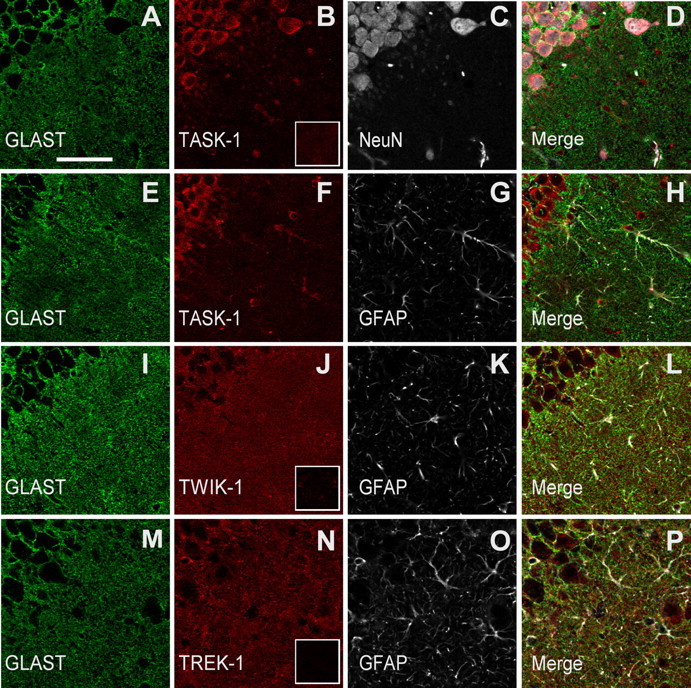
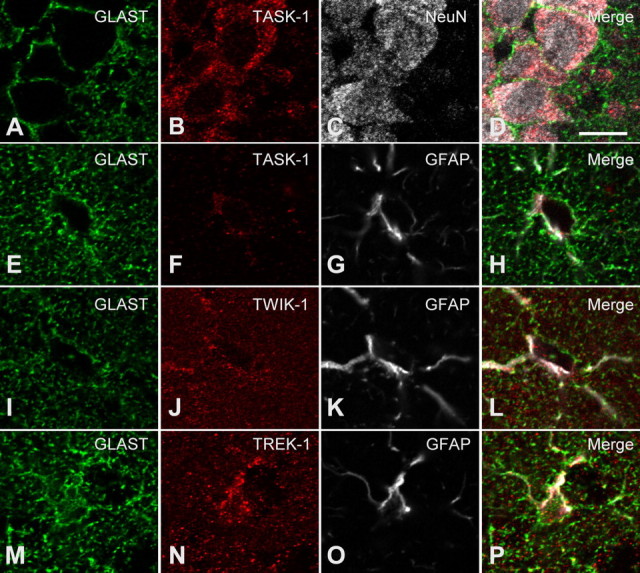
Expression of a linear current-voltage (I-V) relationship (passive) K(+) membrane conductance is a hallmark of mature hippocampal astrocytes. However, the molecular identifications of the K(+) channels underlying this passive conductance remain unknown. We provide the following evidence supporting significant contribution of the two-pore domain K(+) channel (K(2P)) isoforms, TWIK-1 and TREK-1, to this conductance. First, both passive astrocytes and the cloned rat TWIK-1 and TREK-1 channels expressed in CHO cells conduct significant amounts of Cs(+) currents, but vary in their relative P(Cs)/P(K) permeability, 0.43, 0.10, and 0.05, respectively. Second, quinine, which potently inhibited TWIK-1 (IC(50) = 85 microm) and TREK-1 (IC(50) = 41 microm) currents, also inhibited astrocytic passive conductance by 58% at a concentration of 200 microm. Third, a moderate sensitivity of passive conductance to low extracellular pH (6.0) supports a combined expression of acid-insensitive TREK-1, and to a lesser extent, acid-sensitive TWIK-1. Fourth, the astrocyte passive conductance showed low sensitivity to extracellular Ba(2+), and extracellular Ba(2+) blocked TWIK-1 channels at an IC(50) of 960 microm and had no effect on TREK-1 channels. Finally, an immunocytochemical study showed colocalization of TWIK-1 and TREK-1 proteins with the astrocytic markers GLAST and GFAP in rat hippocampal stratum radiatum. In contrast, another K(2P) isoform TASK-1 was mainly colocalized with the neuronal marker NeuN in hippocampal pyramidal neurons and was expressed at a much lower level in astrocytes. These results support TWIK-1 and TREK-1 as being the major components of the long-sought K(+) channels underlying the passive conductance of mature hippocampal astrocytes.
Figures
References
-
- Amzica F, Massimini M. Glial and neuronal interactions during slow wave and paroxysmal activities in the neocortex. Cereb Cortex. 2002;12:1101–1113. - PubMed
-
- Bergles DE, Jahr CE. Synaptic activation of glutamate transporters in hippocampal astrocytes. Neuron. 1997;19:1297–1308. - PubMed
-
- Cahoy JD, Emery B, Kaushal A, Foo LC, Zamanian JL, Christopherson KS, Xing Y, Lubischer JL, Krieg PA, Krupenko SA, Thompson WJ, Barres BA. A transcriptome database for astrocytes, neurons, and oligodendrocytes: a new resource for understanding brain development and function. J Neurosci. 2008;28:264–278. - PMC - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous