

Helical extension of the neuronal SNARE complex into the membrane
- PMID: 19571812
- PMCID: PMC3108252
- DOI: 10.1038/nature08156
Helical extension of the neuronal SNARE complex into the membrane
Abstract
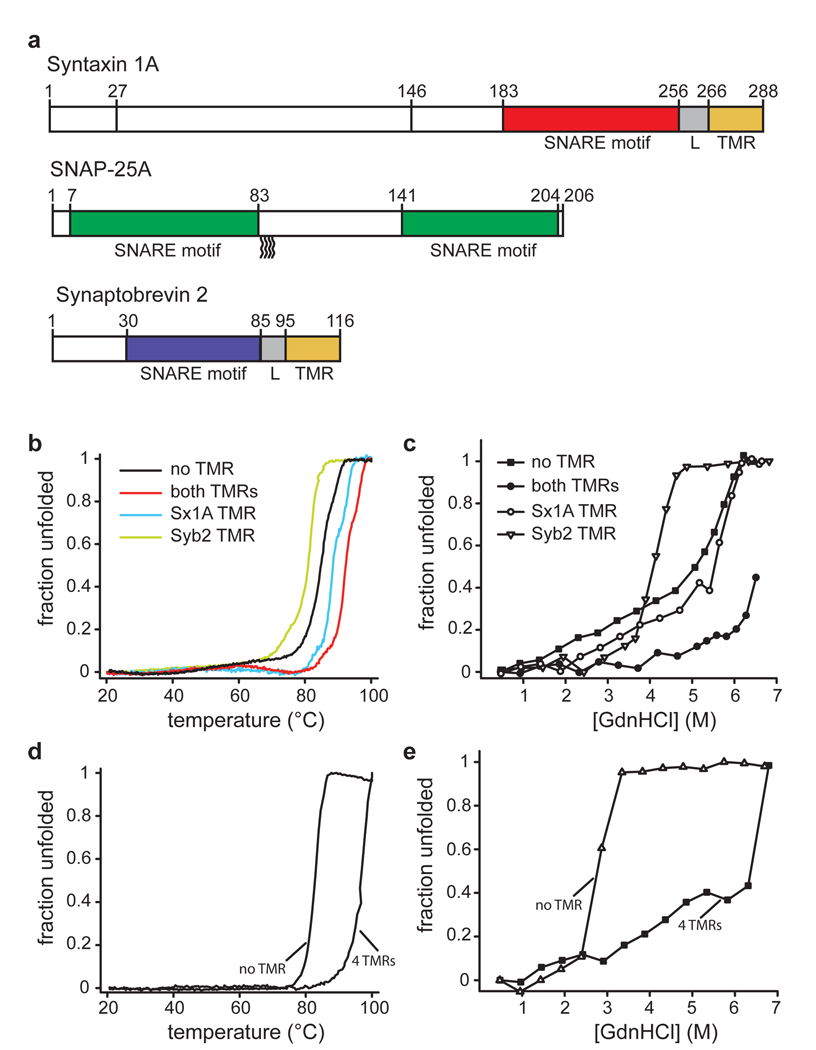
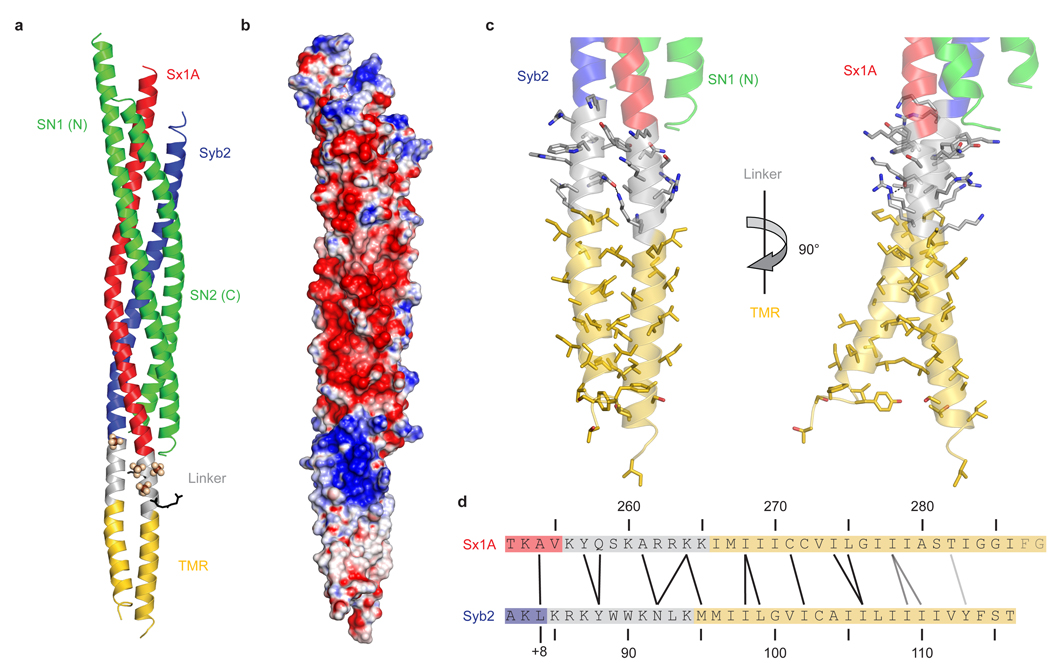
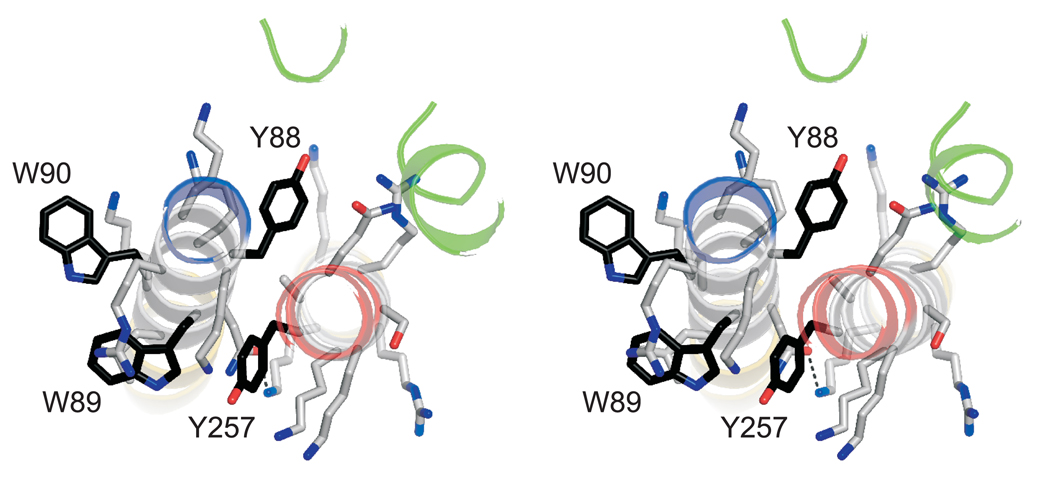
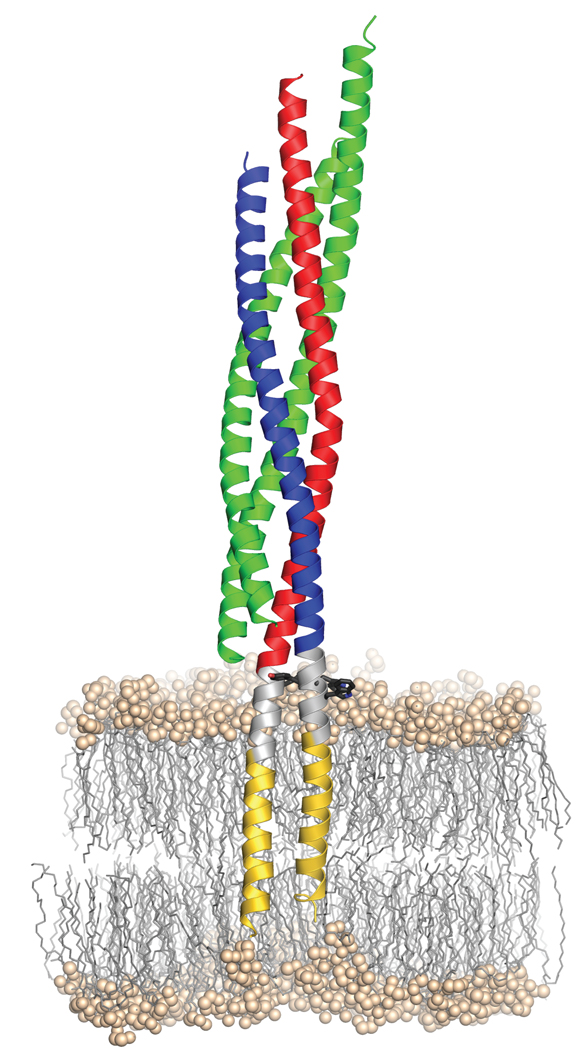
Neurotransmission relies on synaptic vesicles fusing with the membrane of nerve cells to release their neurotransmitter content into the synaptic cleft, a process requiring the assembly of several members of the SNARE (soluble N-ethylmaleimide-sensitive factor attachment protein receptor) family. SNAREs represent an evolutionarily conserved protein family that mediates membrane fusion in the secretory and endocytic pathways of eukaryotic cells. On membrane contact, these proteins assemble in trans between the membranes as a bundle of four alpha-helices, with the energy released during assembly being thought to drive fusion. However, it is unclear how the energy is transferred to the membranes and whether assembly is conformationally linked to fusion. Here, we report the X-ray structure of the neuronal SNARE complex, consisting of rat syntaxin 1A, SNAP-25 and synaptobrevin 2, with the carboxy-terminal linkers and transmembrane regions at 3.4 A resolution. The structure shows that assembly proceeds beyond the already known core SNARE complex, resulting in a continuous helical bundle that is further stabilized by side-chain interactions in the linker region. Our results suggest that the final phase of SNARE assembly is directly coupled to membrane merger.
Figures
References
-
- Martens S, McMahon HT. Mechanisms of membrane fusion: disparate players and common principles. Nat Rev Mol Cell Biol. 2008;9:543–556. - PubMed
-
- Jahn R, Scheller RH. SNAREs--engines for membrane fusion. Nat Rev Mol Cell Biol. 2006;7:631–643. - PubMed
-
- Pelham HR, Banfield DK, Lewis MJ. SNAREs involved in traffic through the Golgi complex. Cold Spring Harb Symp Quant Biol. 1995;60:105–111. - PubMed
-
- Hanson PI, Heuser JE, Jahn R. Neurotransmitter release - four years of SNARE complexes. Curr Opin Neurobiol. 1997;7:310–315. - PubMed
References for Full Methods
-
- Fasshauer D, Eliason WK, Brunger AT, Jahn R. Identification of a minimal core of the synaptic SNARE complex sufficient for reversible assembly and disassembly. Biochemistry. 1998;37:10354–10362. - PubMed
-
- Pobbati AV, et al. Structural basis for the inhibitory role of tomosyn in exocytosis. J Biol Chem. 2004;279:47192–47200. - PubMed
-
- Fasshauer D, et al. Mixed and non-cognate SNARE complexes. Characterization of assembly and biophysical properties. J Biol Chem. 1999;274:15440–15446. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
