

Telomerase modulates Wnt signalling by association with target gene chromatin
- PMID: 19571879
- PMCID: PMC4349391
- DOI: 10.1038/nature08137
Telomerase modulates Wnt signalling by association with target gene chromatin
Abstract
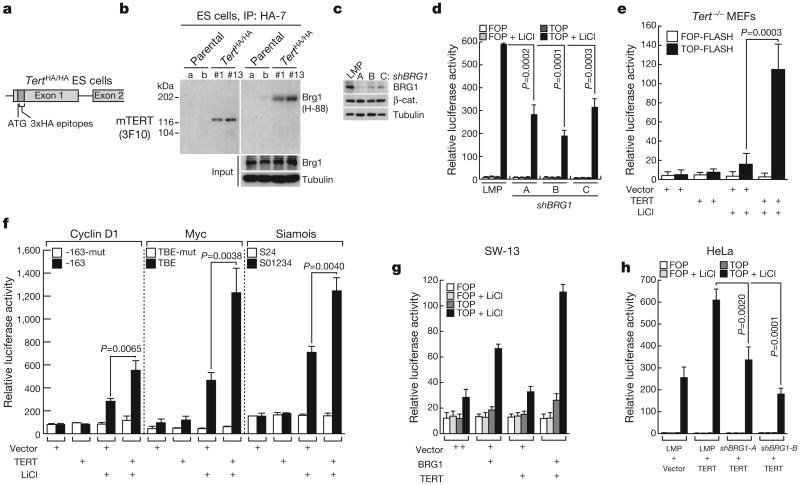
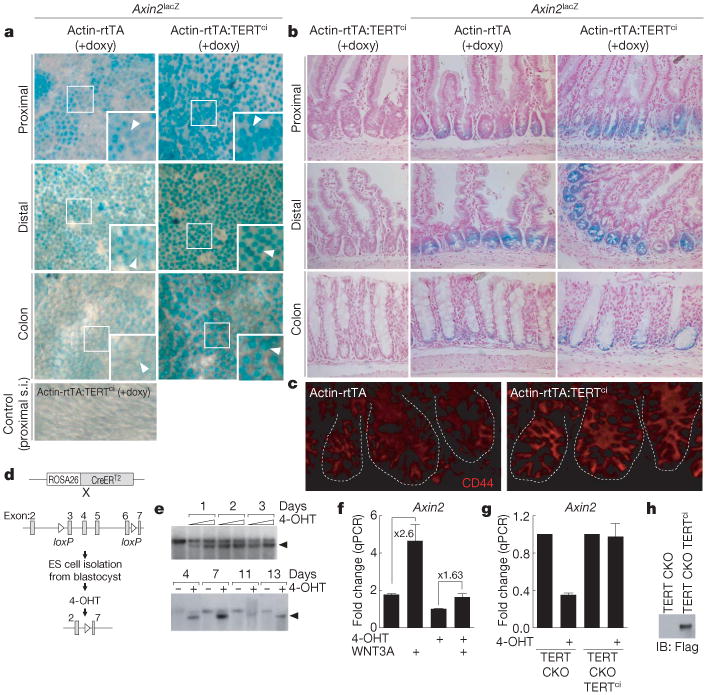
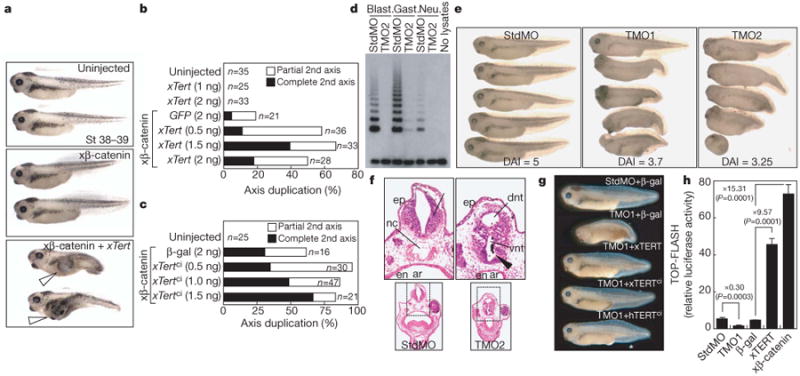
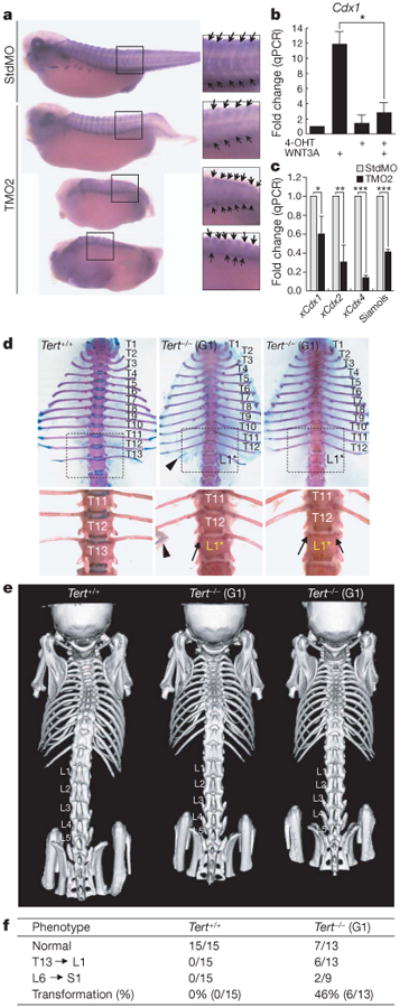
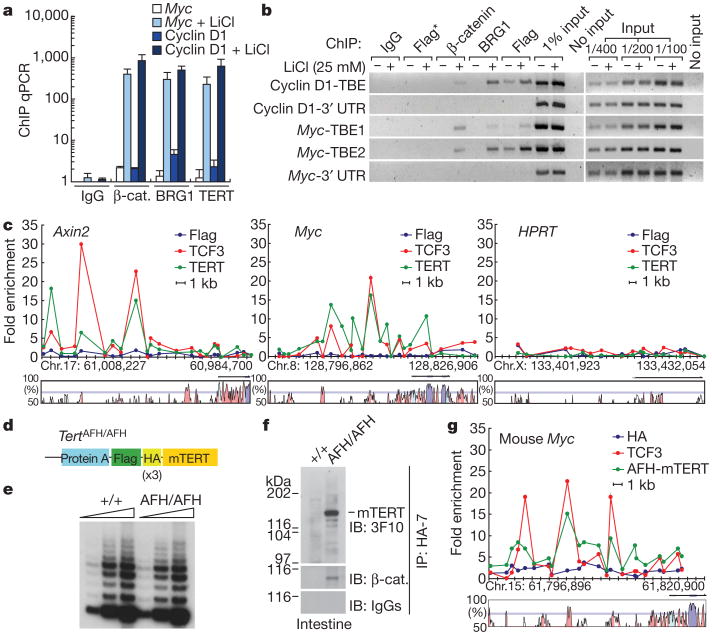
Stem cells are controlled, in part, by genetic pathways frequently dysregulated during human tumorigenesis. Either stimulation of Wnt/beta-catenin signalling or overexpression of telomerase is sufficient to activate quiescent epidermal stem cells in vivo, although the mechanisms by which telomerase exerts these effects are not understood. Here we show that telomerase directly modulates Wnt/beta-catenin signalling by serving as a cofactor in a beta-catenin transcriptional complex. The telomerase protein component TERT (telomerase reverse transcriptase) interacts with BRG1 (also called SMARCA4), a SWI/SNF-related chromatin remodelling protein, and activates Wnt-dependent reporters in cultured cells and in vivo. TERT serves an essential role in formation of the anterior-posterior axis in Xenopus laevis embryos, and this defect in Wnt signalling manifests as homeotic transformations in the vertebrae of Tert(-/-) mice. Chromatin immunoprecipitation of the endogenous TERT protein from mouse gastrointestinal tract shows that TERT physically occupies gene promoters of Wnt-dependent genes. These data reveal an unanticipated role for telomerase as a transcriptional modulator of the Wnt/beta-catenin signalling pathway.
Figures
Comment in
-
Cell biology: The not-so-odd couple.Nature. 2009 Jul 2;460(7251):44-5. doi: 10.1038/460044a. Nature. 2009. PMID: 19571874 No abstract available.
References
-
- Reya T, Clevers H. Wnt signalling in stem cells and cancer. Nature. 2005;434:843–850. - PubMed
-
- Gat U, DasGupta R, Degenstein L, Fuchs E. De Novo hair follicle morphogenesis and hair tumors in mice expressing a truncated β-catenin in skin. Cell. 1998;95:605–614. - PubMed
-
- Lo Celso C, Prowse DM, Watt FM. Transient activation of β-catenin signalling in adult mouse epidermis is sufficient to induce new hair follicles but continuous activation is required to maintain hair follicle tumours. Development. 2004;131:1787–1799. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
