

5'-triphosphate RNA requires base-paired structures to activate antiviral signaling via RIG-I
- PMID: 19574455
- PMCID: PMC2705279
- DOI: 10.1073/pnas.0900971106
5'-triphosphate RNA requires base-paired structures to activate antiviral signaling via RIG-I
Abstract
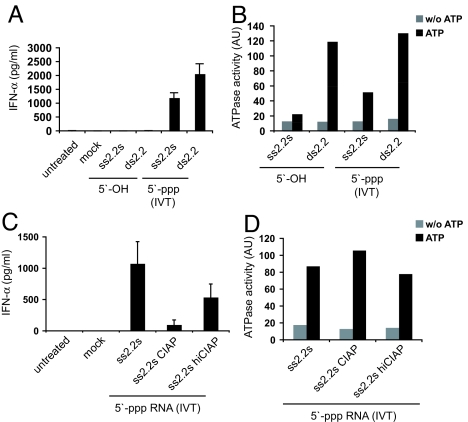
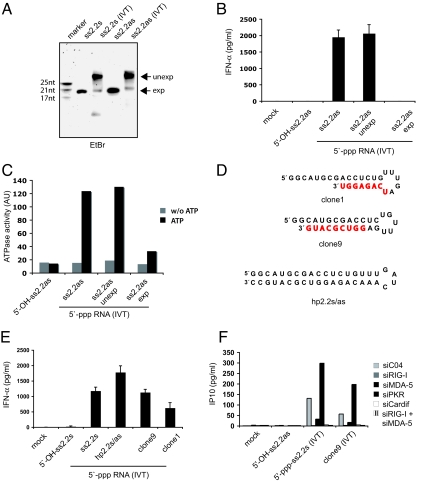
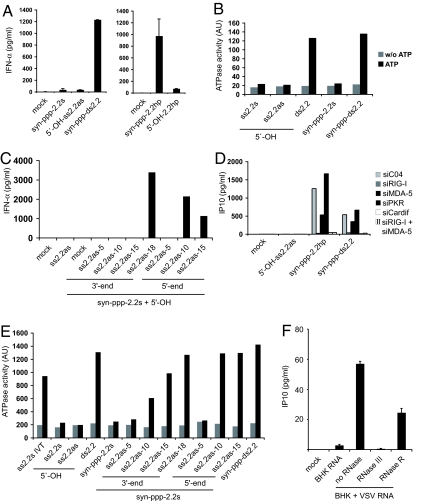
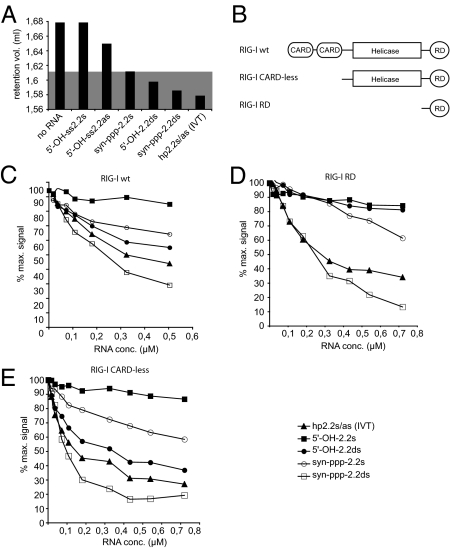
The ATPase retinoid acid-inducible gene (RIG)-I senses viral RNA in the cytoplasm of infected cells and subsequently activates cellular antiviral defense mechanisms. RIG-I recognizes molecular structures that discriminate viral from host RNA. Here, we show that RIG-I ligands require base-paired structures in conjunction with a free 5'-triphosphate to trigger antiviral signaling. Hitherto unavailable chemically synthesized 5'-triphosphate RNA ligands do not trigger RIG-I-dependent IFN production in cells, and they are unable to trigger the ATPase activity of RIG-I without a base-paired stretch. Consistently, immunostimulatory RNA from cells infected with a virus recognized by RIG-I is sensitive to double-strand, but not single-strand, specific RNases. In vitro, base-paired stretches and the 5'-triphosphate bind to distinct sites of RIG-I and synergize to trigger the induction of signaling competent RIG-I multimers. Strengthening our model of a bipartite molecular pattern for RIG-I activation, we show that the activity of supposedly "single-stranded" 5'-triphosphate RNAs generated by in vitro transcription depends on extended and base-paired by-products inadvertently, but commonly, produced by this method. Together, our findings accurately define a minimal molecular pattern sufficient to activate RIG-I that can be found in viral genomes or transcripts.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Akira S, Uematsu S, Takeuchi O. Pathogen recognition and innate immunity. Cell. 2006;124:783–801. - PubMed
-
- Rothenfusser S, et al. The RNA helicase Lgp2 inhibits TLR-independent sensing of viral replication by retinoic acid-inducible gene-I. J Immunol. 2005;175:5260–5268. - PubMed
-
- Yoneyama M, et al. The RNA helicase RIG-I has an essential function in double-stranded RNA-induced innate antiviral responses. Nat Immunol. 2004;5:730–737. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
