

Quantitative time-lapse fluorescence microscopy in single cells
- PMID: 19575655
- PMCID: PMC3137897
- DOI: 10.1146/annurev.cellbio.042308.113408
Quantitative time-lapse fluorescence microscopy in single cells
Abstract
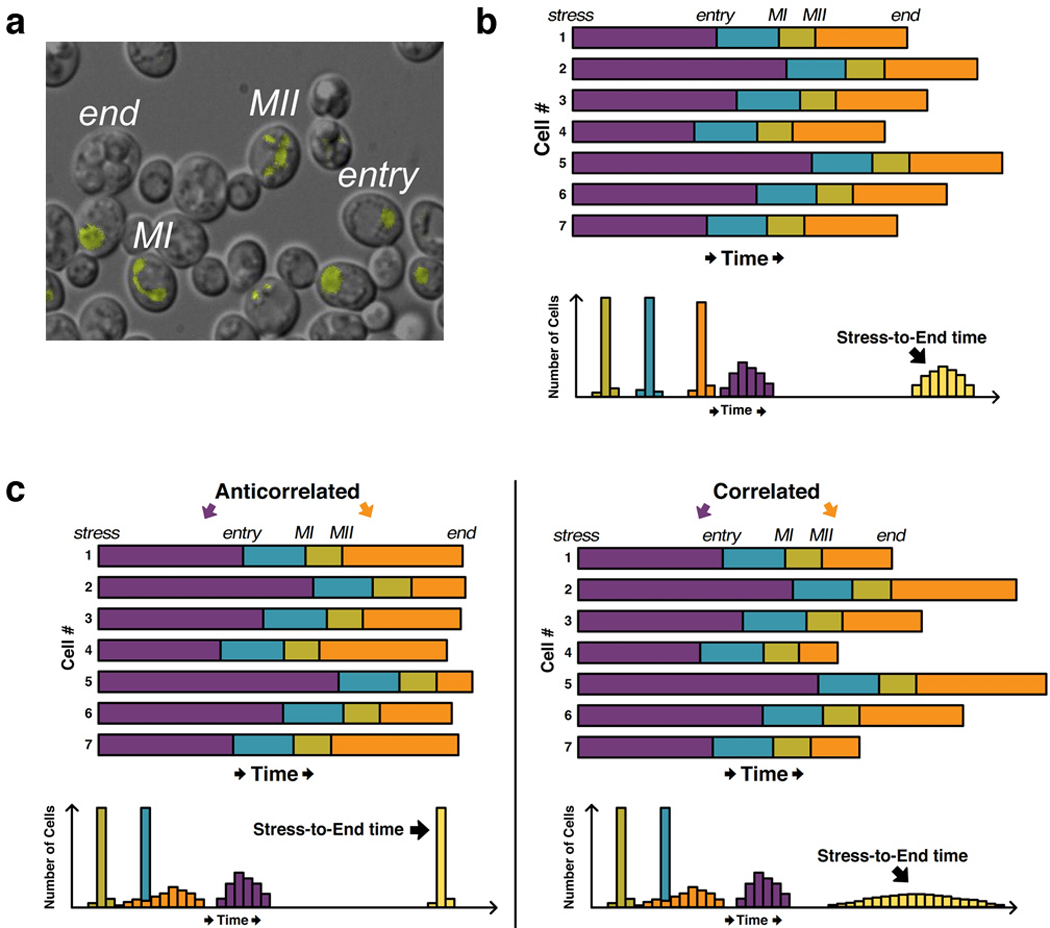
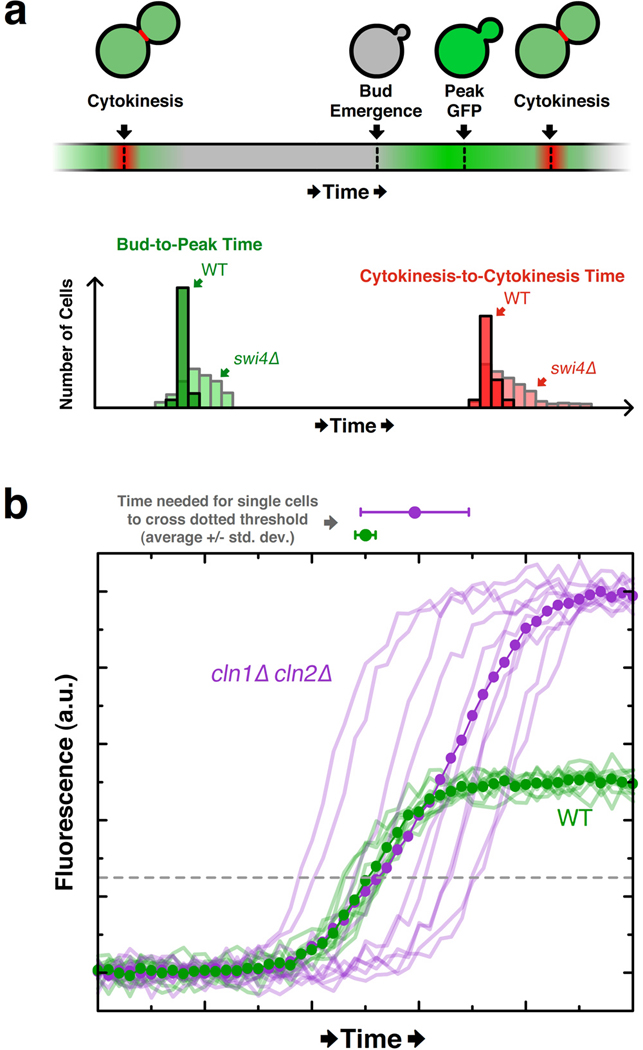
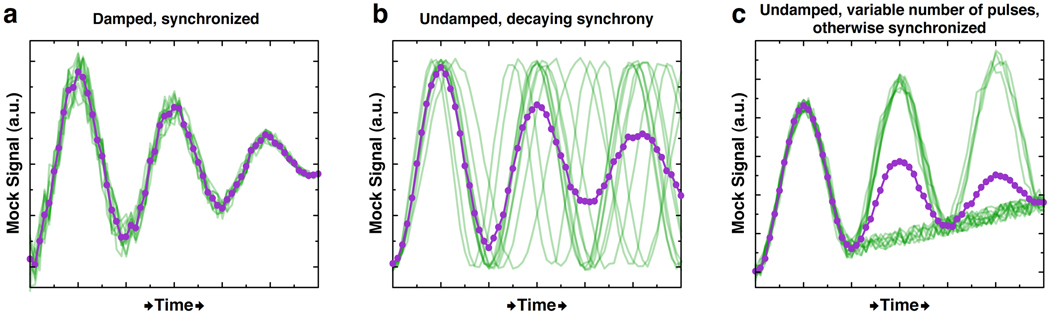
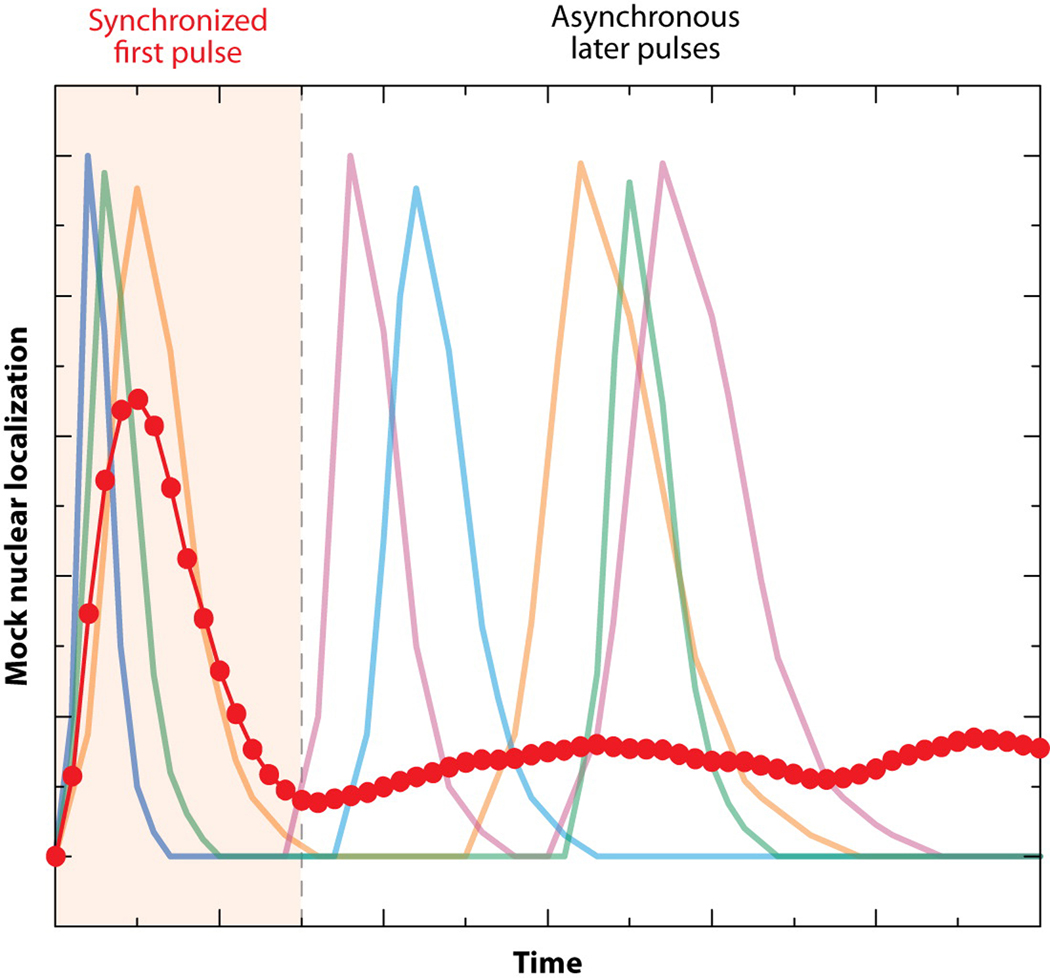
The cloning of green fluorescent protein (GFP) 15 years ago revolutionized cell biology by permitting visualization of a wide range of molecular mechanisms within living cells. Though initially used to make largely qualitative assessments of protein levels and localizations, fluorescence microscopy has since evolved to become highly quantitative and high-throughput. Computational image analysis has catalyzed this evolution, enabling rapid and automated processing of large datasets. Here, we review studies that combine time-lapse fluorescence microscopy and automated image analysis to investigate dynamic events at the single-cell level. We highlight examples where single-cell analysis provides unique mechanistic insights into cellular processes that cannot be otherwise resolved in bulk assays. Additionally, we discuss studies where quantitative microscopy facilitates the assembly of detailed 4D lineages in developing organisms. Finally, we describe recent advances in imaging technology, focusing especially on platforms that allow the simultaneous perturbation and quantitative monitoring of biological systems.
Figures

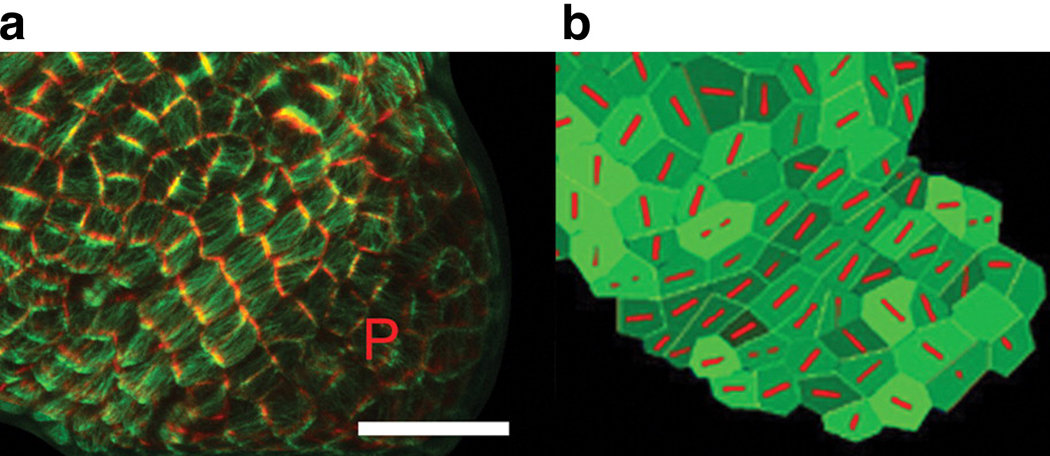
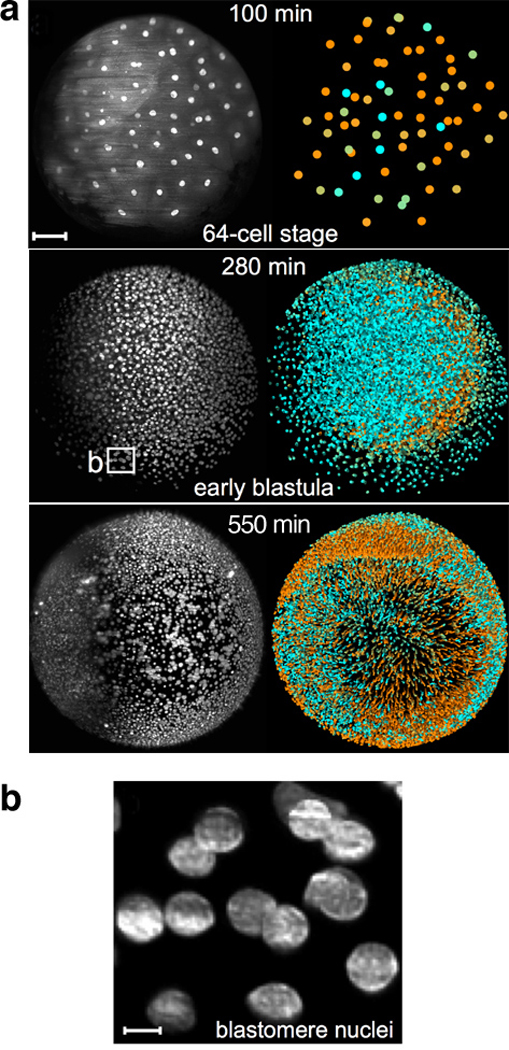
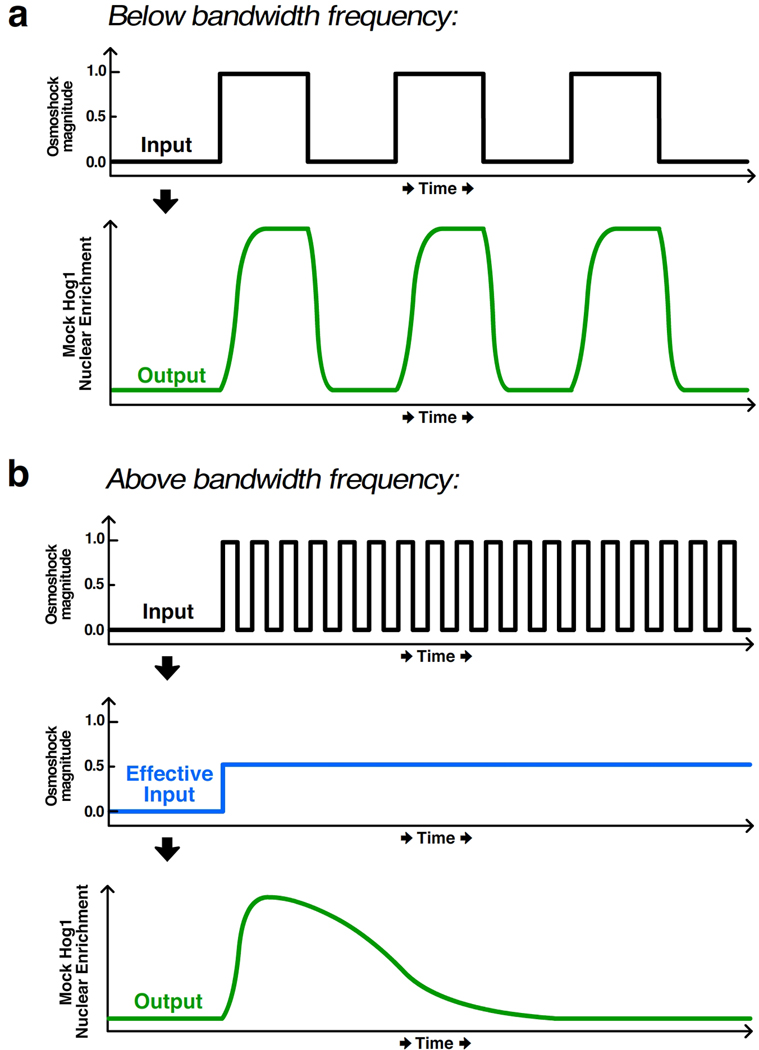
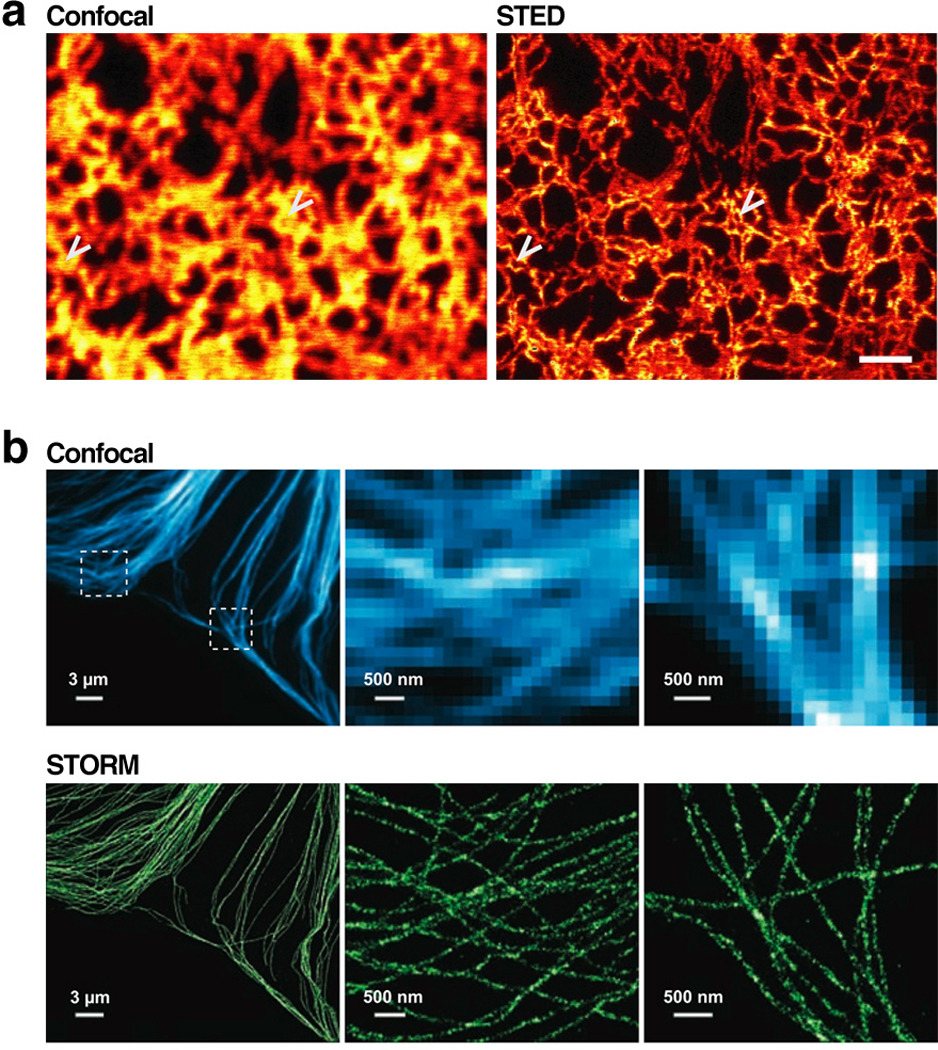
Similar articles
-
Automated quantitative live cell fluorescence microscopy.Cold Spring Harb Perspect Biol. 2010 Aug;2(8):a000455. doi: 10.1101/cshperspect.a000455. Epub 2010 Jun 30. Cold Spring Harb Perspect Biol. 2010. PMID: 20591990 Free PMC article. Review.
-
High-throughput RNAi screening by time-lapse imaging of live human cells.Nat Methods. 2006 May;3(5):385-90. doi: 10.1038/nmeth876. Nat Methods. 2006. PMID: 16628209
-
An automatic method for robust and fast cell detection in bright field images from high-throughput microscopy.BMC Bioinformatics. 2013 Oct 4;14:297. doi: 10.1186/1471-2105-14-297. BMC Bioinformatics. 2013. PMID: 24090363 Free PMC article.
-
A faster, high resolution, mtPA-GFP-based mitochondrial fusion assay acquiring kinetic data of multiple cells in parallel using confocal microscopy.J Vis Exp. 2012 Jul 20;(65):e3991. doi: 10.3791/3991. J Vis Exp. 2012. PMID: 22847388 Free PMC article.
-
Quantitative motion analysis and visualization of cellular structures.Methods. 2003 Jan;29(1):3-13. doi: 10.1016/s1046-2023(02)00287-6. Methods. 2003. PMID: 12543067 Review.
Cited by
-
Spatial mapping of protein composition and tissue organization: a primer for multiplexed antibody-based imaging.Nat Methods. 2022 Mar;19(3):284-295. doi: 10.1038/s41592-021-01316-y. Epub 2021 Nov 22. Nat Methods. 2022. PMID: 34811556 Free PMC article. Review.
-
Microfluidic picoliter bioreactor for microbial single-cell analysis: fabrication, system setup, and operation.J Vis Exp. 2013 Dec 6;(82):50560. doi: 10.3791/50560. J Vis Exp. 2013. PMID: 24336165 Free PMC article.
-
A high-throughput microscopy method for single-cell analysis of event-time correlations in nanoparticle-induced cell death.Commun Biol. 2019 Jan 24;2:35. doi: 10.1038/s42003-019-0282-0. eCollection 2019. Commun Biol. 2019. PMID: 30701200 Free PMC article.
-
Establishment of Signaling Interactions with Cellular Resolution for Every Cell Cycle of Embryogenesis.Genetics. 2018 May;209(1):37-49. doi: 10.1534/genetics.118.300820. Epub 2018 Mar 22. Genetics. 2018. PMID: 29567658 Free PMC article.
-
Gradient matching accelerates mixed-effects inference for biochemical networks.Bioinformatics. 2025 Mar 29;41(4):btaf154. doi: 10.1093/bioinformatics/btaf154. Bioinformatics. 2025. PMID: 40199819 Free PMC article.
References
-
- Ai HW, Shaner NC, Cheng Z, Tsien RY, Campbell RE. Exploration of new chromophore structures leads to the identification of improved blue fluorescent proteins. Biochemistry. 2007;46:5904–5910. - PubMed
-
- Ando R, Mizuno H, Miyawaki A. Regulated fast nucleocytoplasmic shuttling observed by reversible protein highlighting. Science. 2004;306:1370–1373. - PubMed
-
- Balagaddé FK, You L, Hansen CL, Arnold FH, Quake SR. Long-term monitoring of bacteria undergoing programmed population control in a microchemostat. Science. 2005;309:137–140. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
