

Recognition of 5' triphosphate by RIG-I helicase requires short blunt double-stranded RNA as contained in panhandle of negative-strand virus
- PMID: 19576794
- PMCID: PMC2824854
- DOI: 10.1016/j.immuni.2009.05.008
Recognition of 5' triphosphate by RIG-I helicase requires short blunt double-stranded RNA as contained in panhandle of negative-strand virus
Abstract
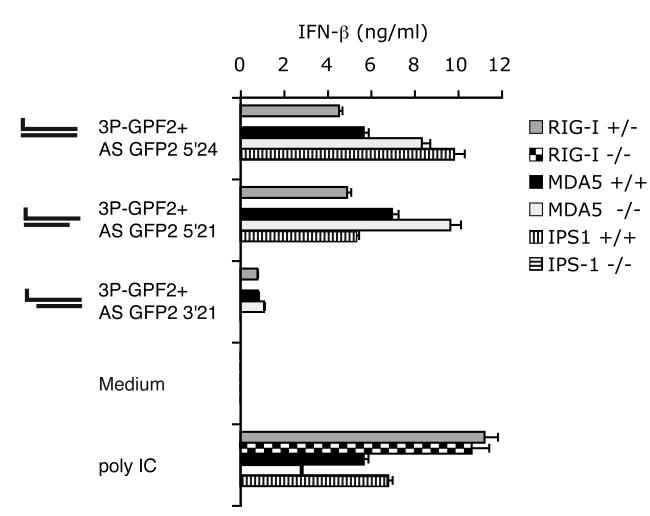
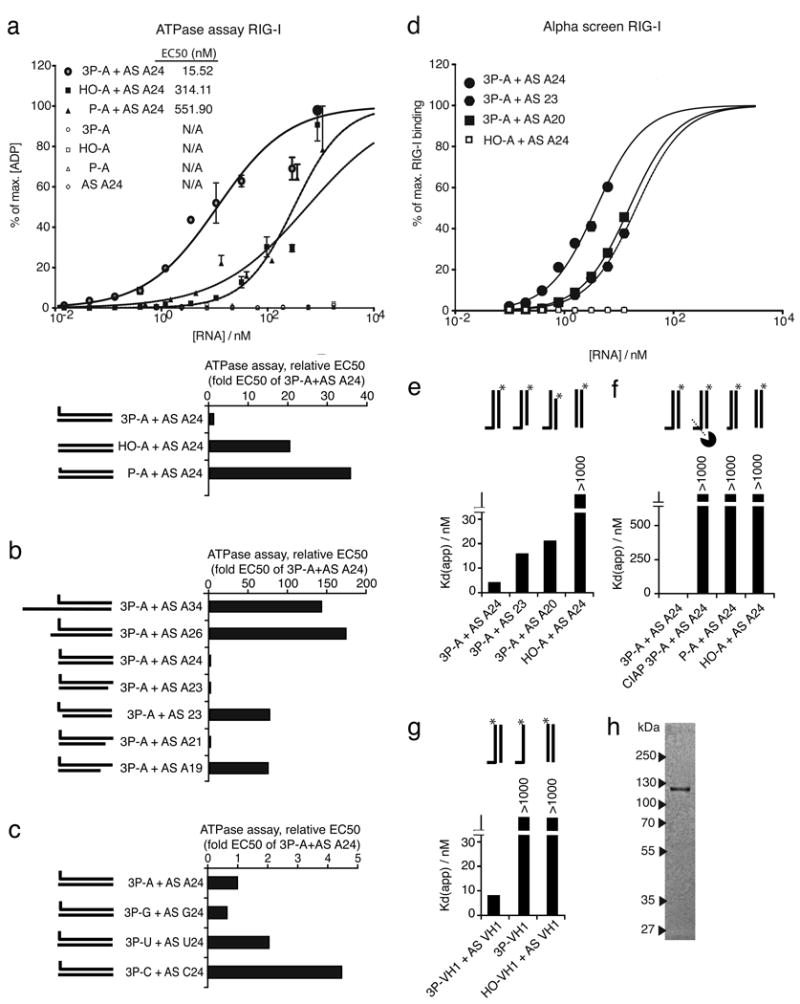
Antiviral immunity is triggered by immunorecognition of viral nucleic acids. The cytosolic helicase RIG-I is a key sensor of viral infections and is activated by RNA containing a triphosphate at the 5' end. The exact structure of RNA activating RIG-I remains controversial. Here, we established a chemical approach for 5' triphosphate oligoribonucleotide synthesis and found that synthetic single-stranded 5' triphosphate oligoribonucleotides were unable to bind and activate RIG-I. Conversely, the addition of the synthetic complementary strand resulted in optimal binding and activation of RIG-I. Short double-strand conformation with base pairing of the nucleoside carrying the 5' triphosphate was required. RIG-I activation was impaired by a 3' overhang at the 5' triphosphate end. These results define the structure of RNA for full RIG-I activation and explain how RIG-I detects negative-strand RNA viruses that lack long double-stranded RNA but do contain blunt short double-stranded 5' triphosphate RNA in the panhandle region of their single-stranded genome.
Figures
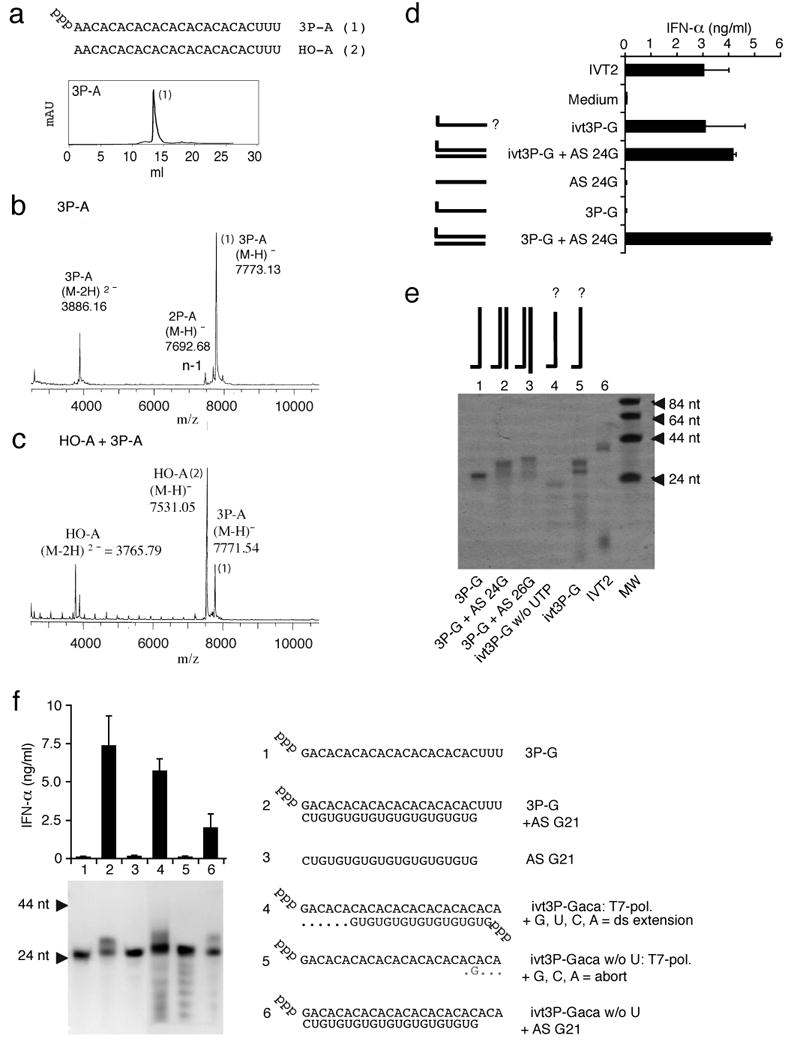
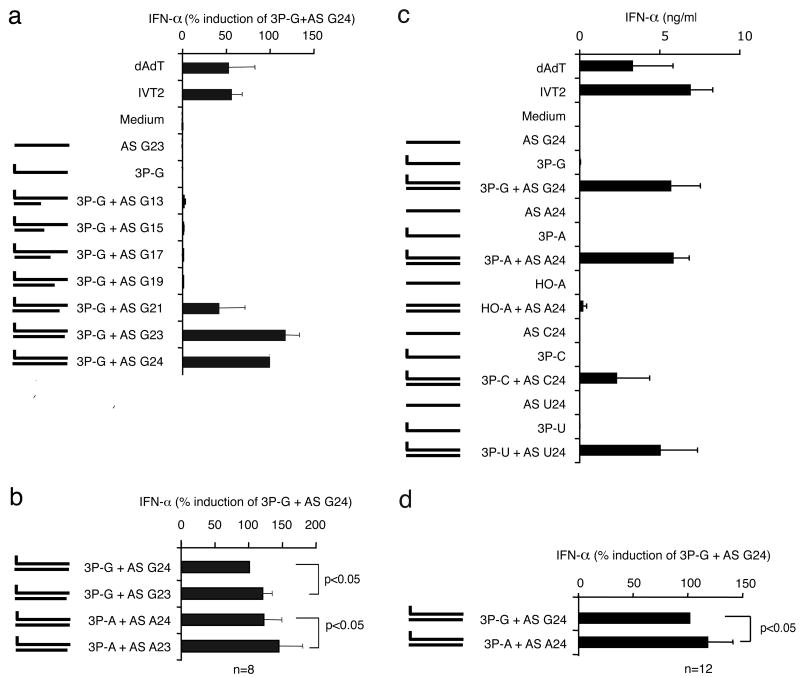
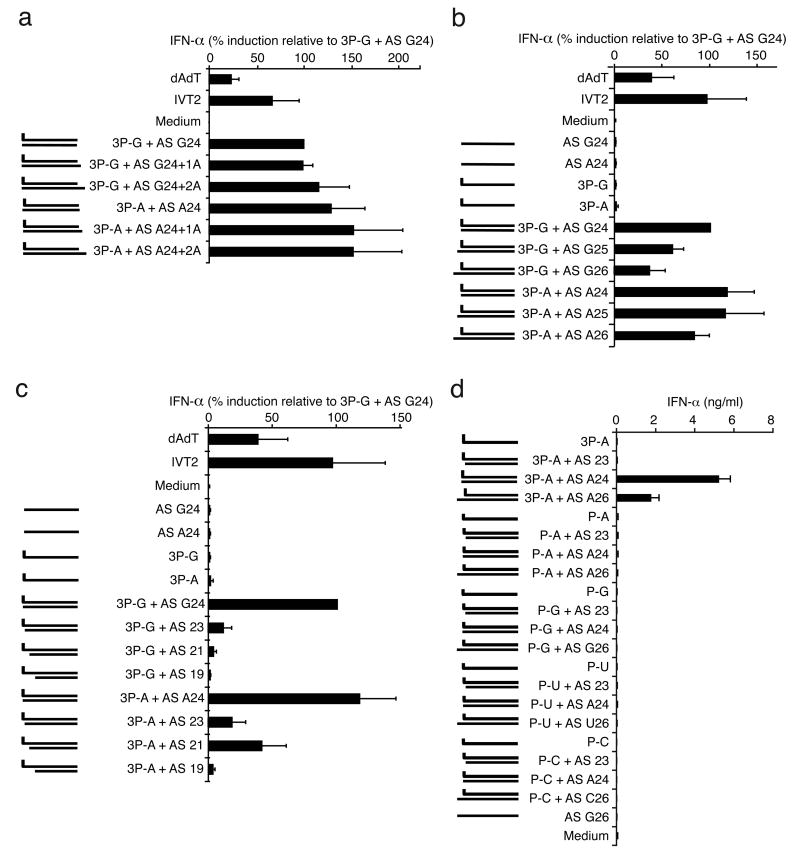
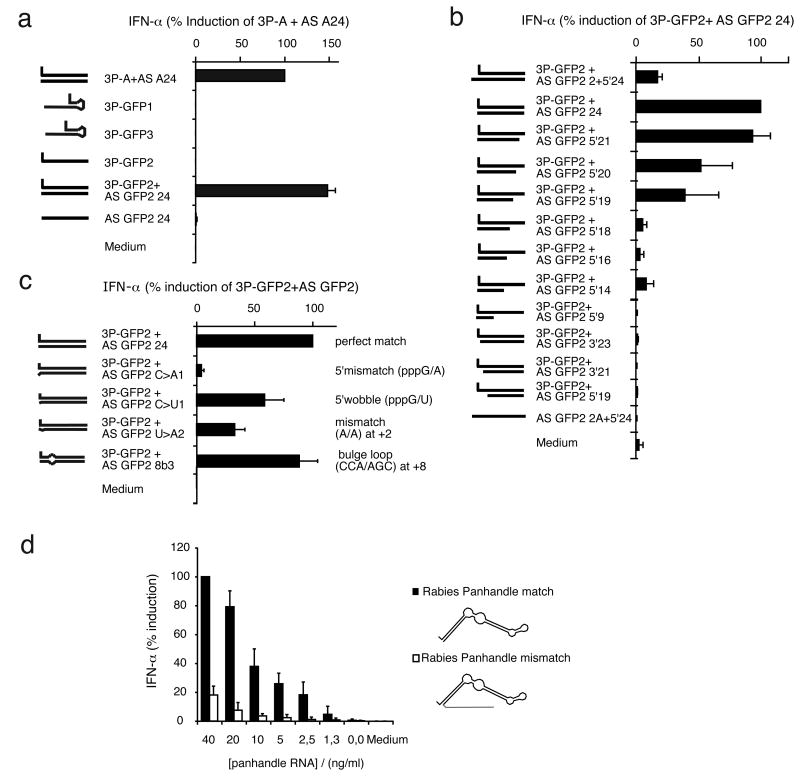

Comment in
-
A nonself RNA pattern: tri-p to panhandle.Immunity. 2009 Jul 17;31(1):4-5. doi: 10.1016/j.immuni.2009.06.014. Immunity. 2009. PMID: 19604485
References
-
- Cui S, Eisenacher K, Kirchhofer A, Brzozka K, Lammens A, Lammens K, Fujita T, Conzelmann KK, Krug A, Hopfner KP. The C-terminal regulatory domain is the RNA 5′-triphosphate sensor of RIG-I. Mol Cell. 2008;29:169–179. - PubMed
-
- Fire A, Xu S, Montgomery MK, Kostas SA, Driver SE, Mello CC. Potent and specific genetic interference by double-stranded RNA in Caenorhabditis elegans. Nature. 1998;391:806–811. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
