

Thiolutin inhibits endothelial cell adhesion by perturbing Hsp27 interactions with components of the actin and intermediate filament cytoskeleton
- PMID: 19579057
- PMCID: PMC2866983
- DOI: 10.1007/s12192-009-0130-0
Thiolutin inhibits endothelial cell adhesion by perturbing Hsp27 interactions with components of the actin and intermediate filament cytoskeleton
Abstract
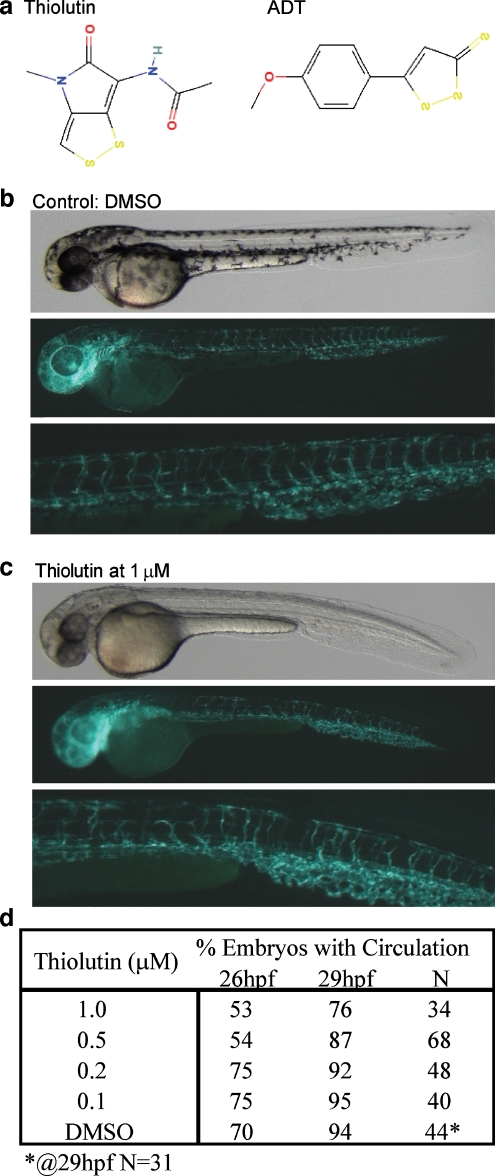
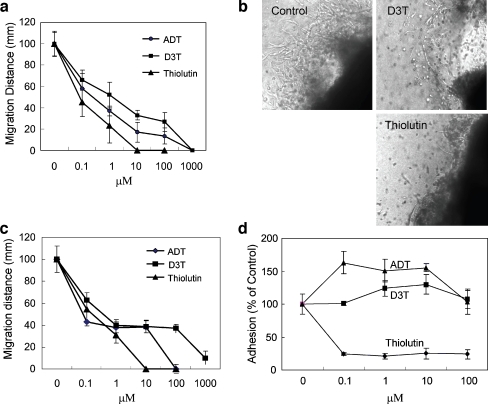
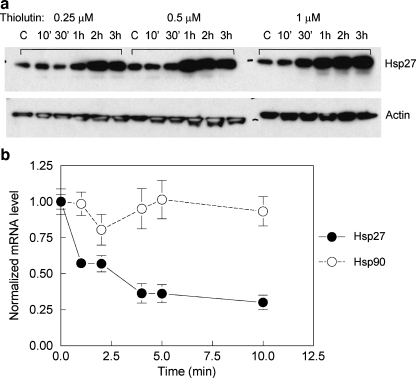
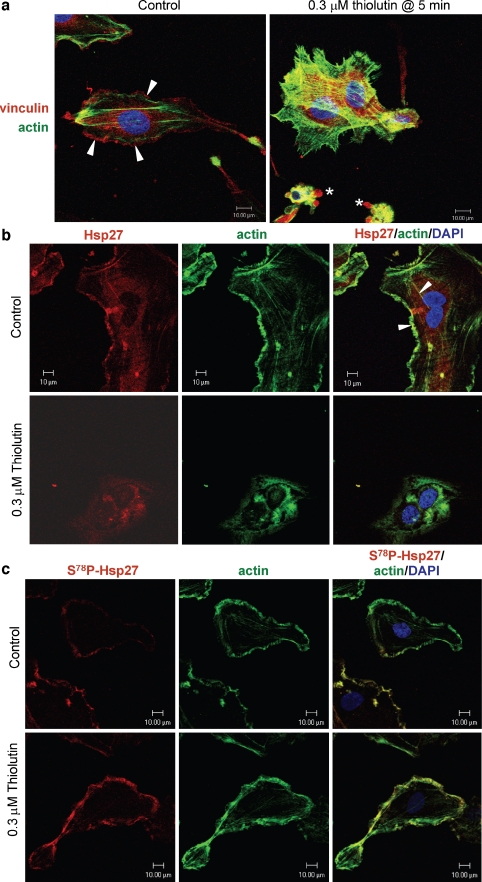
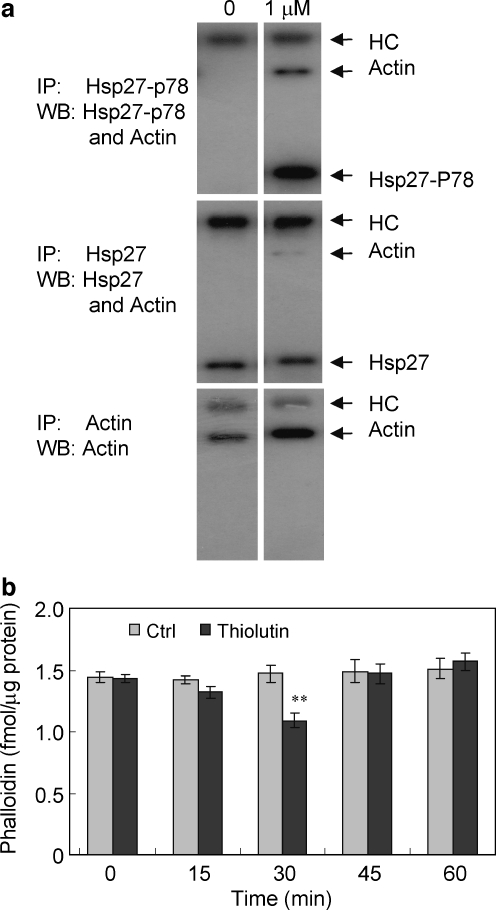
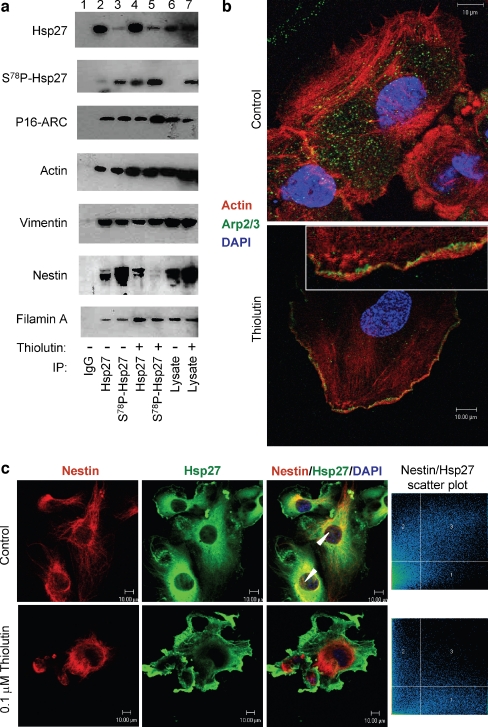
Thiolutin is a dithiole synthesized by Streptomyces sp. that inhibits endothelial cell adhesion and tumor growth. We show here that thiolutin potently inhibits developmental angiogenesis in zebrafish and vascular outgrowth from tissue explants in 3D cultures. Thiolutin is a potent and selective inhibitor of endothelial cell adhesion accompanied by rapid induction of HSPB1 (Hsp27) phosphorylation. The inhibitory effects of thiolutin on endothelial cell adhesion are transient, potentially due to a compensatory increase in Hsp27 protein levels. Accordingly, heat shock induction of Hsp27 limits the anti-adhesive activity of thiolutin. Thiolutin treatment results in loss of actin stress fibers, increased cortical actin as cells retract, and decreased cellular F-actin. Mass spectrometric analysis of Hsp27 binding partners following immunoaffinity purification identified several regulatory components of the actin cytoskeleton that associate with Hsp27 in a thiolutin-sensitive manner including several components of the Arp2/3 complex. Among these, ArpC1a is a direct binding partner of Hsp27. Thiolutin treatment induces peripheral localization of phosphorylated Hsp27 and Arp2/3. Hsp27 also associates with the intermediate filament components vimentin and nestin. Thiolutin treatment specifically ablates Hsp27 interaction with nestin and collapses nestin filaments. These results provide new mechanistic insights into regulation of cell adhesion and cytoskeletal dynamics by Hsp27.
Figures

References
-
- Bitar KN. HSP27 phosphorylation and interaction with actin-myosin in smooth muscle contraction. Am J Physiol Gastrointest Liver Physiol. 2002;282:G894–903. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
