

Caffeine suppresses amyloid-beta levels in plasma and brain of Alzheimer's disease transgenic mice
- PMID: 19581723
- PMCID: PMC3746074
- DOI: 10.3233/JAD-2009-1071
Caffeine suppresses amyloid-beta levels in plasma and brain of Alzheimer's disease transgenic mice
Erratum in
- J Alzheimers Dis. 2009 Nov;18(3):727. Wang, Lilly [corrected to Wang, Li].
Abstract
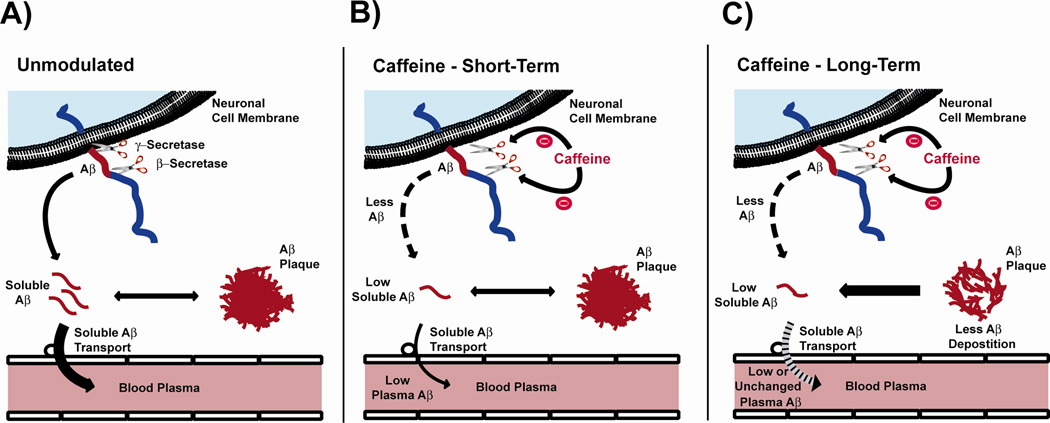
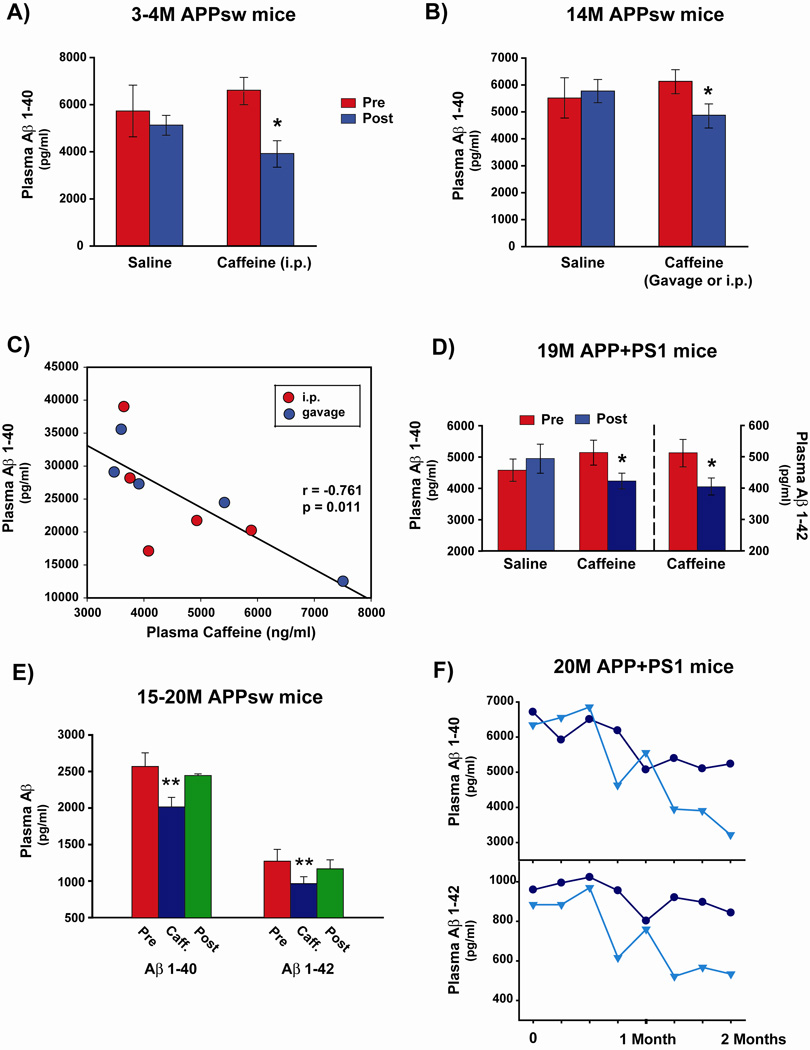
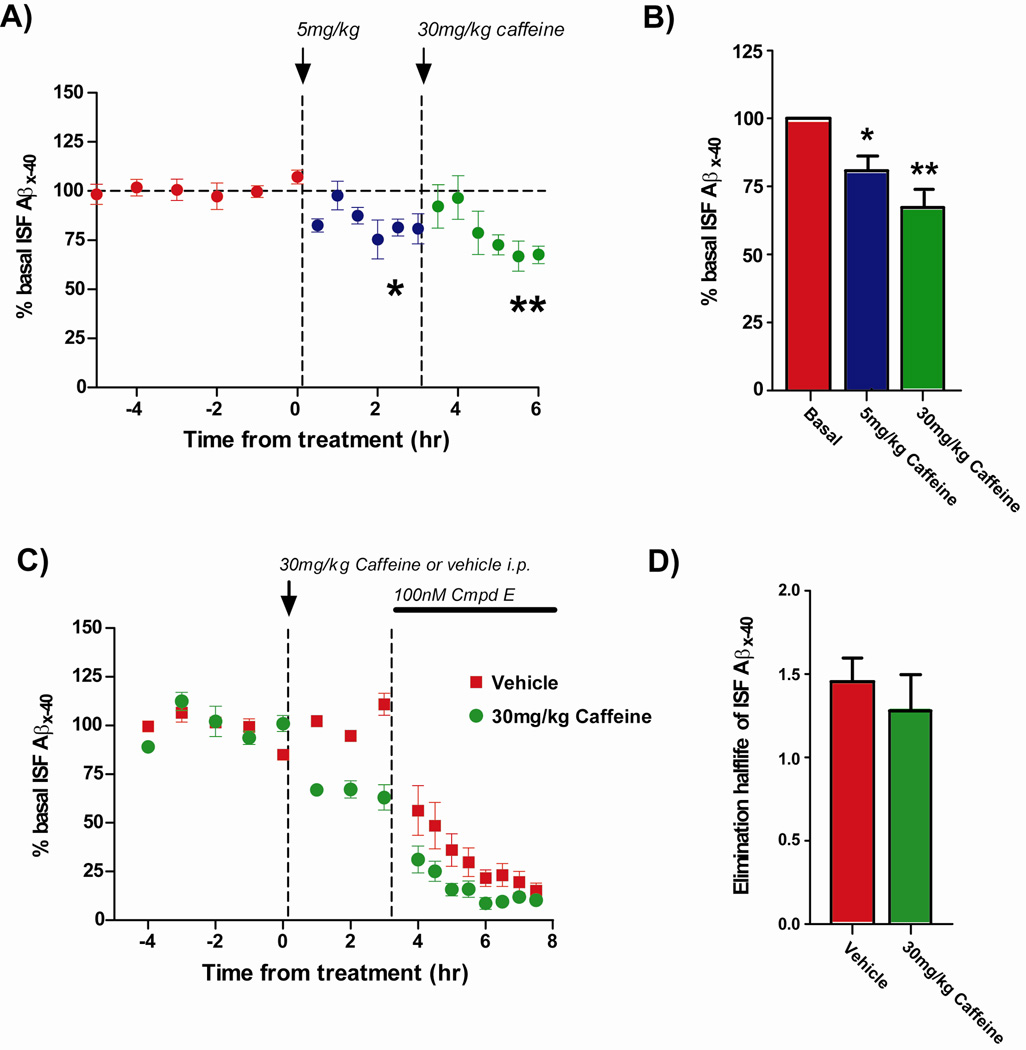
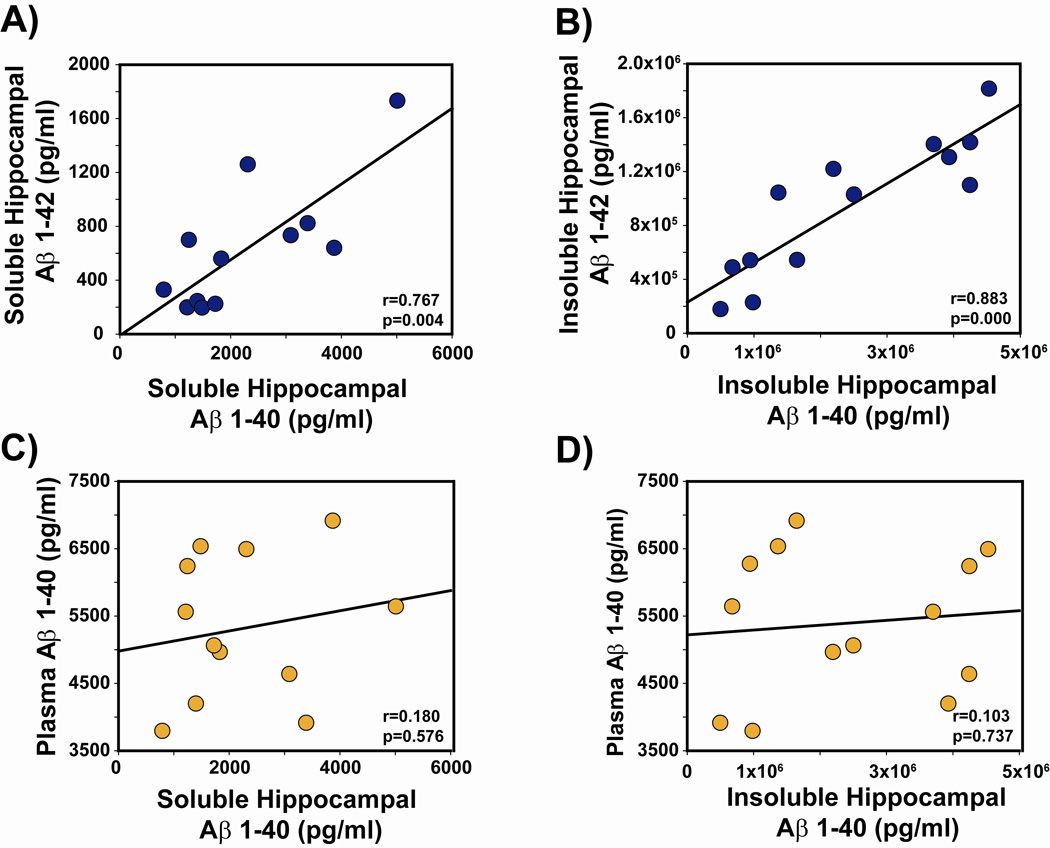
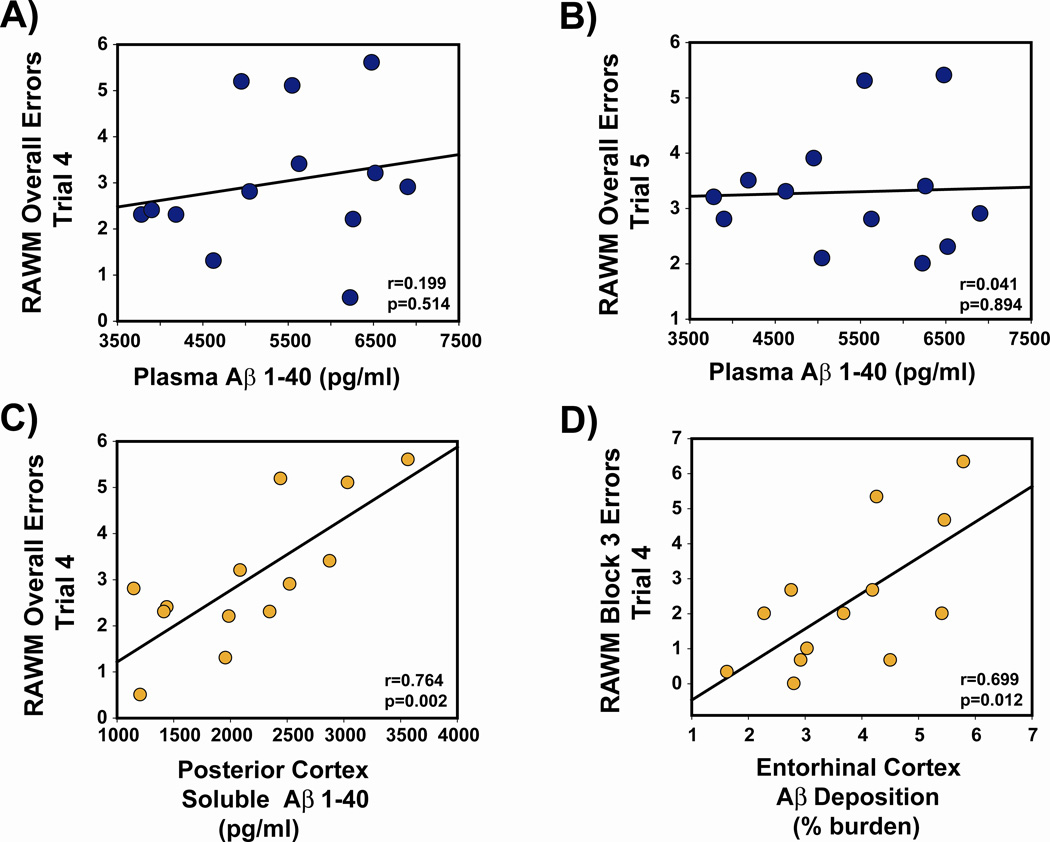
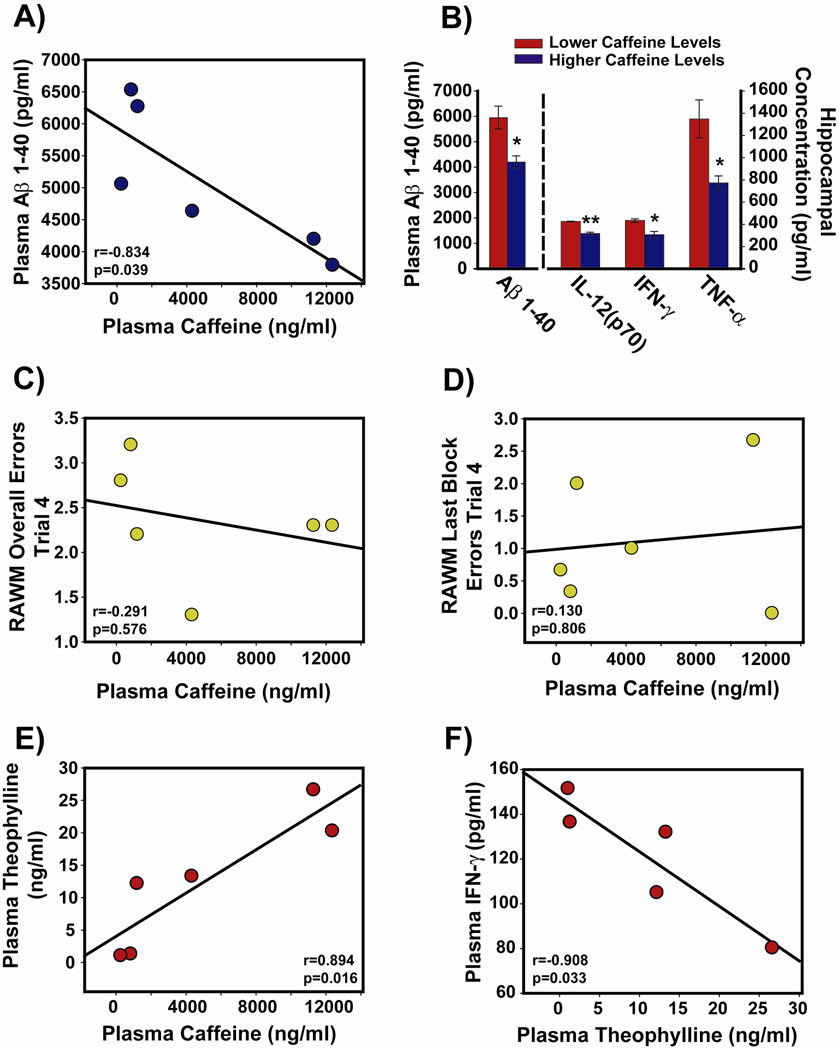
Recent epidemiologic studies suggest that caffeine may be protective against Alzheimer's disease (AD). Supportive of this premise, our previous studies have shown that moderate caffeine administration protects/restores cognitive function and suppresses brain amyloid-beta (Abeta) production in AD transgenic mice. In the present study, we report that acute caffeine administration to both young adult and aged AD transgenic mice rapidly reduces Abeta levels in both brain interstitial fluid and plasma without affecting Abeta elimination. Long-term oral caffeine treatment to aged AD mice provided not only sustained reductions in plasma Abeta, but also decreases in both soluble and deposited Abeta in hippocampus and cortex. Irrespective of caffeine treatment, plasma Abeta levels did not correlate with brain Abeta levels or with cognitive performance in individual aged AD mice. Although higher plasma caffeine levels were strongly associated with lower plasma Abeta1-40 levels in aged AD mice, plasma caffeine levels were also not linked to cognitive performance. Plasma caffeine and theophylline levels were tightly correlated, both being associated with reduced inflammatory cytokine levels in hippocampus. Our conclusion is two-fold: first, that both plasma and brain Abeta levels are reduced by acute or chronic caffeine administration in several AD transgenic lines and ages, indicating a therapeutic value of caffeine against AD; and second, that plasma Abeta levels are not an accurate index of brain Abeta levels/deposition or cognitive performance in aged AD mice.
Figures

Similar articles
-
Caffeine reverses cognitive impairment and decreases brain amyloid-beta levels in aged Alzheimer's disease mice.J Alzheimers Dis. 2009;17(3):661-80. doi: 10.3233/JAD-2009-1087. J Alzheimers Dis. 2009. PMID: 19581722
-
Caffeine synergizes with another coffee component to increase plasma GCSF: linkage to cognitive benefits in Alzheimer's mice.J Alzheimers Dis. 2011;25(2):323-35. doi: 10.3233/JAD-2011-110110. J Alzheimers Dis. 2011. PMID: 21422521
-
Caffeine protects Alzheimer's mice against cognitive impairment and reduces brain beta-amyloid production.Neuroscience. 2006 Nov 3;142(4):941-52. doi: 10.1016/j.neuroscience.2006.07.021. Epub 2006 Aug 28. Neuroscience. 2006. PMID: 16938404
-
Caffeine and coffee as therapeutics against Alzheimer's disease.J Alzheimers Dis. 2010;20 Suppl 1:S117-26. doi: 10.3233/JAD-2010-091249. J Alzheimers Dis. 2010. PMID: 20182037 Review.
-
Caffeine protects against disruptions of the blood-brain barrier in animal models of Alzheimer's and Parkinson's diseases.J Alzheimers Dis. 2010;20 Suppl 1(Suppl 1):S127-41. doi: 10.3233/JAD-2010-1376. J Alzheimers Dis. 2010. PMID: 20164568 Free PMC article. Review.
Cited by
-
Diet in the Prevention of Alzheimer's Disease: Current Knowledge and Future Research Requirements.Nutrients. 2022 Oct 30;14(21):4564. doi: 10.3390/nu14214564. Nutrients. 2022. PMID: 36364826 Free PMC article. Review.
-
Association between coffee and tea consumption and the risk of dementia in individuals with hypertension: a prospective cohort study.Sci Rep. 2024 Sep 10;14(1):21063. doi: 10.1038/s41598-024-71426-y. Sci Rep. 2024. PMID: 39256489 Free PMC article.
-
Altered motility of plaque-associated microglia in a model of Alzheimer's disease.Neuroscience. 2016 Aug 25;330:410-20. doi: 10.1016/j.neuroscience.2016.05.061. Epub 2016 Jun 7. Neuroscience. 2016. PMID: 27288150 Free PMC article.
-
Pharmacological Effect of Caffeine on Drosophila melanogaster: A Proof-of-Concept in vivo Study for Nootropic Investigation.Arch Razi Inst. 2021 Dec 30;76(6):1645-1654. doi: 10.22092/ARI.2021.356628.1884. eCollection 2021 Dec. Arch Razi Inst. 2021. PMID: 35546991 Free PMC article.
-
Combined long-term enriched environment and caffeine supplementation improve memory function in C57Bl6 mice.Eur Arch Psychiatry Clin Neurosci. 2023 Feb;273(1):269-281. doi: 10.1007/s00406-022-01431-7. Epub 2022 Jun 9. Eur Arch Psychiatry Clin Neurosci. 2023. PMID: 35676374 Free PMC article.
References
-
- Fredholm B, Bättig K, Holmén J, Nehlig A, Zvartau E. Actions of caffeine in the brain with special reference to factors that contribute to its widespread use. Pharmacol Rev. 1999;51:83–133. - PubMed
-
- Higdon J, Frei B. Coffee and health: a review of recent human research. Crit Rev Food Sci Nutr. 2006;46:101–123. - PubMed
-
- Ritchie K, Carrière I, de Mendonca A, Portet F, Dartigues J, Rouaud O, Barberger-Gateau P, Ancelin M. The neuroprotective effects of caffeine: a prospective population study (the Three City Study) Neurology. 2007;69:536–545. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
